- Understand and identify the stages of follicular growth (primordial, primary, secondary, tertiary), as well as the changes that occur in the follicular wall during pregnancy.
- Identify the regional variations in the structure of the oviduct.
- Describe the changes that occur in the ovary and oviduct during the menstrual cycle.
The female reproductive system is composed of highly specialized organs which are in a state of constant change, from the sequential alterations characteristic of each menstrual cycle to the dramatic changes that occur during pregnancy. You will examine the ovary which contains the germ cells or oocytes, the oviduct (Fallopian tube) which receives the ovum at ovulation and conducts it to the uterus where a fertilized egg may implant. At term, the fetus passes through the uterine cervix and the vagina to the external environment.
Slide 239 Ovary, monkey, H&E View Virtual Slide
Slide 269 Ovary, monkey, PAS stain View Virtual Slide
Slide 235 Ovary, human, H&E View Virtual Slide
Slide 234 Ovary, human, H&E View Virtual Slide
Slide 234-1 Ovary, human, H&E View Virtual Slide
Slide 234-2 Ovary, human, trichrome stain View Virtual Slide
Slide 236a Ovary, human, H&E View Virtual Slide
Overview: The ovaries are paired organs situated on either side of the uterus. They are attached on one edge, the hilus, to the broad ligament of the uterus by a fold of peritoneum, the mesovarium. Using slide 239, examine the overall topography of the ovary and note the numerous vessels which enter it via the broad ligament. The inner medulla (present in most slides) is highly vascular and composed of a loose connective tissue core. Examine the outer cortex of the ovary which is composed of stroma and numerous follicles in various stages of development. In slide 239, note the layer of collagenous connective tissue, the tunica albuginea View Image just below the surface epithelium (mesothelium/serosa often misleadingly referred to as "germinal epithelium") that covers the ovary.
{kind=link}
Examine the stroma of the cortex in slide 239 and note the whorls of closely-packed, spindle-shaped fibroblasts. The cortex also contains many oocytes (300,000- 400,000 at birth) embedded in this cortical stroma. Because of the variation in sectioning, age and stage of the cycle, you will have to study several slides in order to study all aspects of follicular development, atresia and corpus luteum formation.
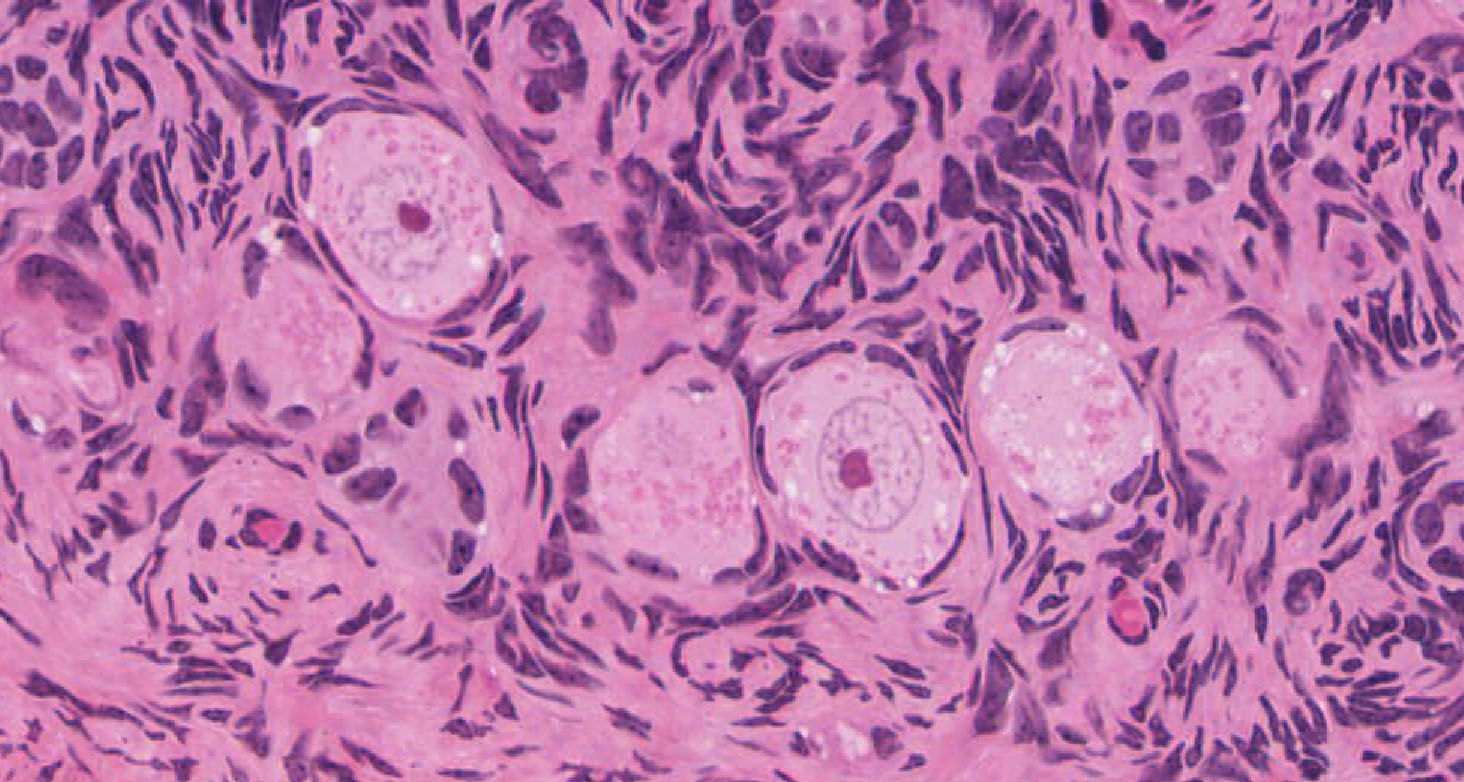
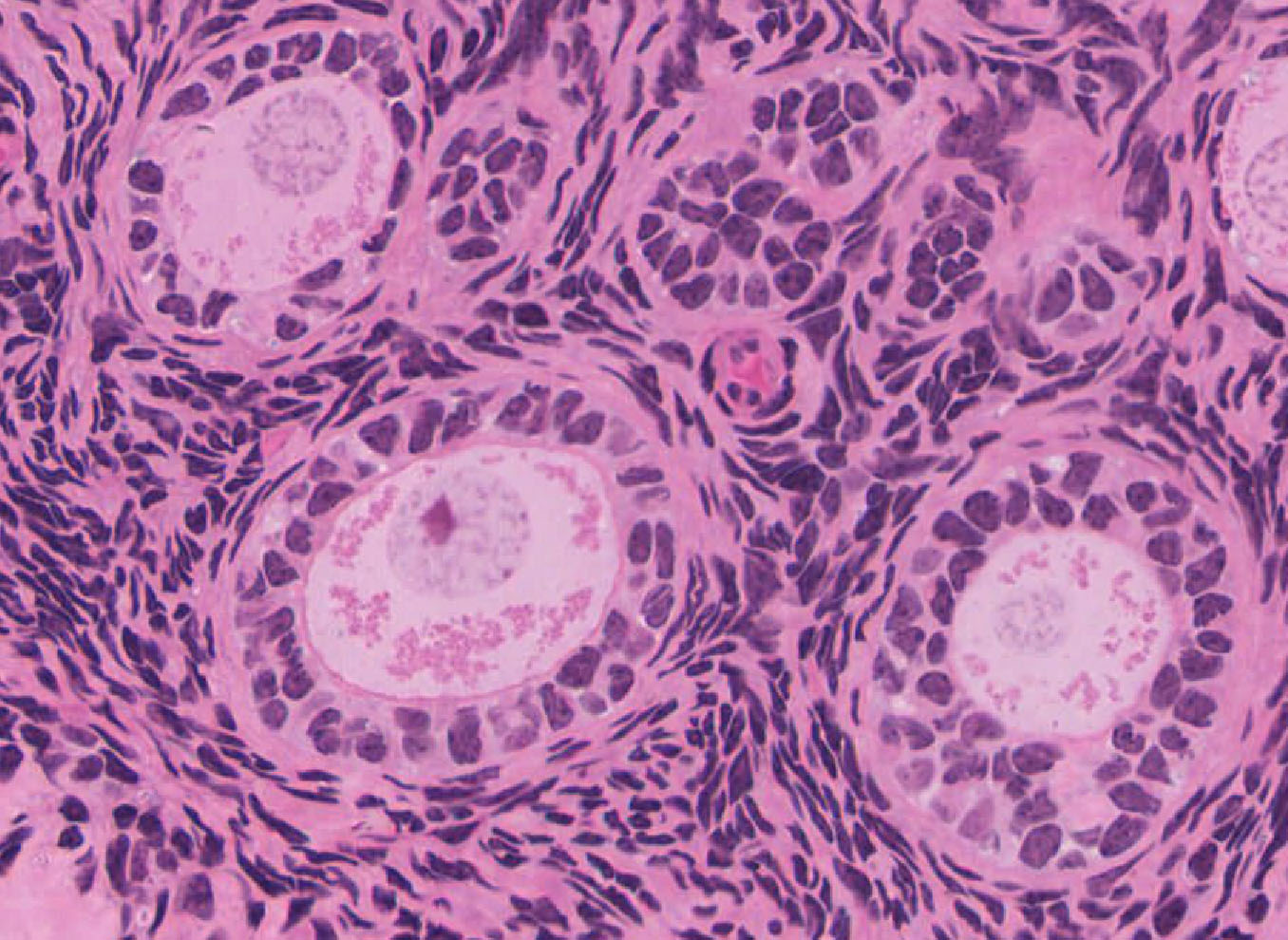
Primordial & Primary Follicles: Examine several primordial follicles View Image using slide 239 or 269 and note that they consist of a large oocyte surrounded by a layer of flattened follicular cells. Next examine the appearance of primary follicles View Image in which the large oocyte is surrounded by a layer of cuboidal follicular cells (also good in slide 238). These follicular cells proliferate to form a loose multi-layer, the granulosa cell layer. A rim of neutral glycoprotein, the zona pellucida (clear zone), surrounds the oocyte separating it from the surrounding granulosa cells. Slide 269 has been stained with PAS, so that carbohydrates and connective tissue are highlighted. Using this slide, examine the zona pellucida View Image of several smaller follicles. Stromal cells form a dense sheath (theca) around the follicle.
{kind=link}
{kind=link}
{kind=link}
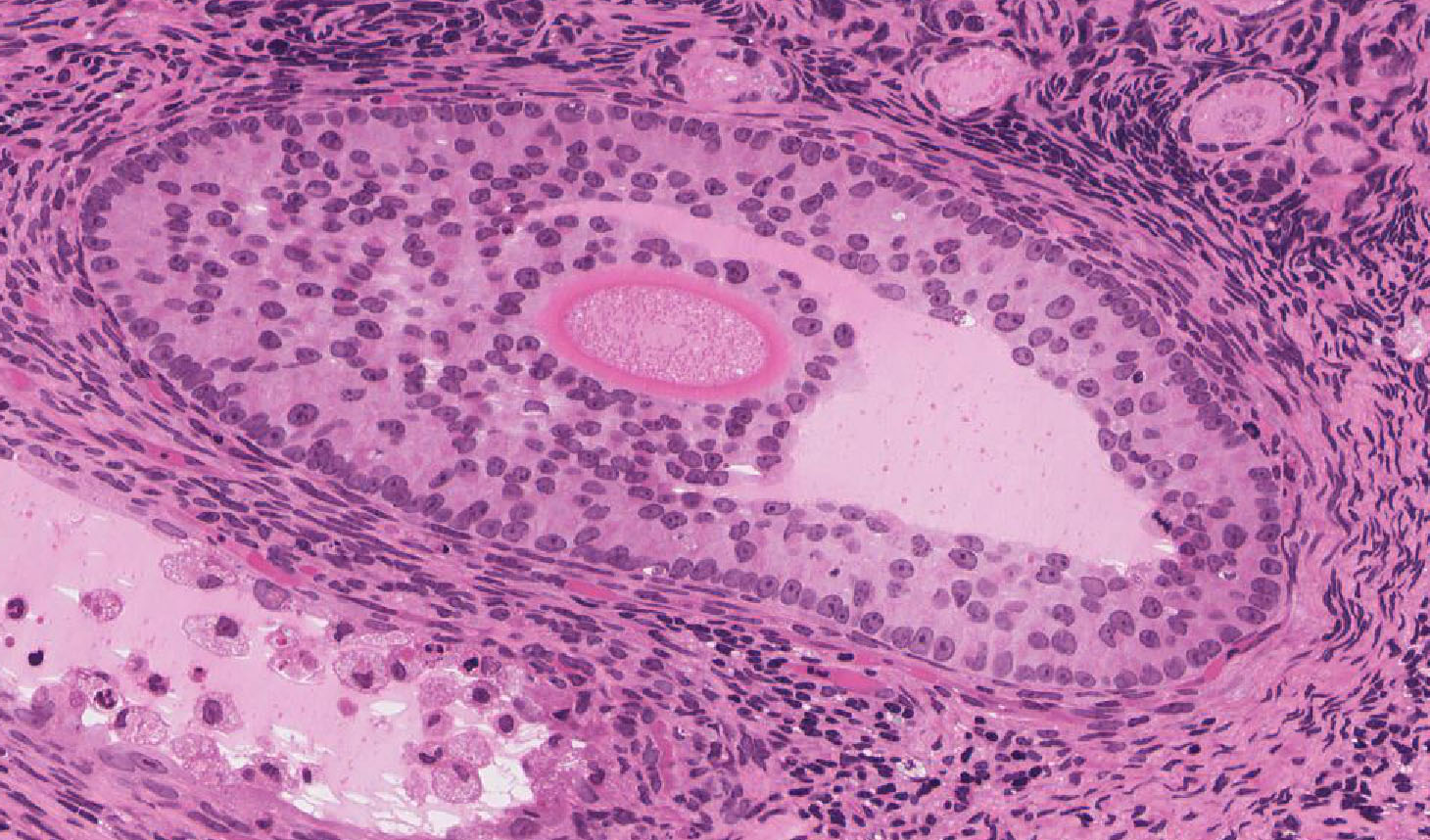
Secondary Follicles: Examine the structure of several secondary follicles View Image and observe that between the stratified granulosa cells there are large lacunae that coalesce to form the follicular antrum. The stromal cells surrounding the follicle have differentiated to form an inner layer (theca interna) of plump cells that secrete steroid precursors and an outer layer (theca externa) composed of concentrically arranged stromal cells that provide support for the developing follicle. Slide 235 also has good theca layers (see below).
{kind=link}
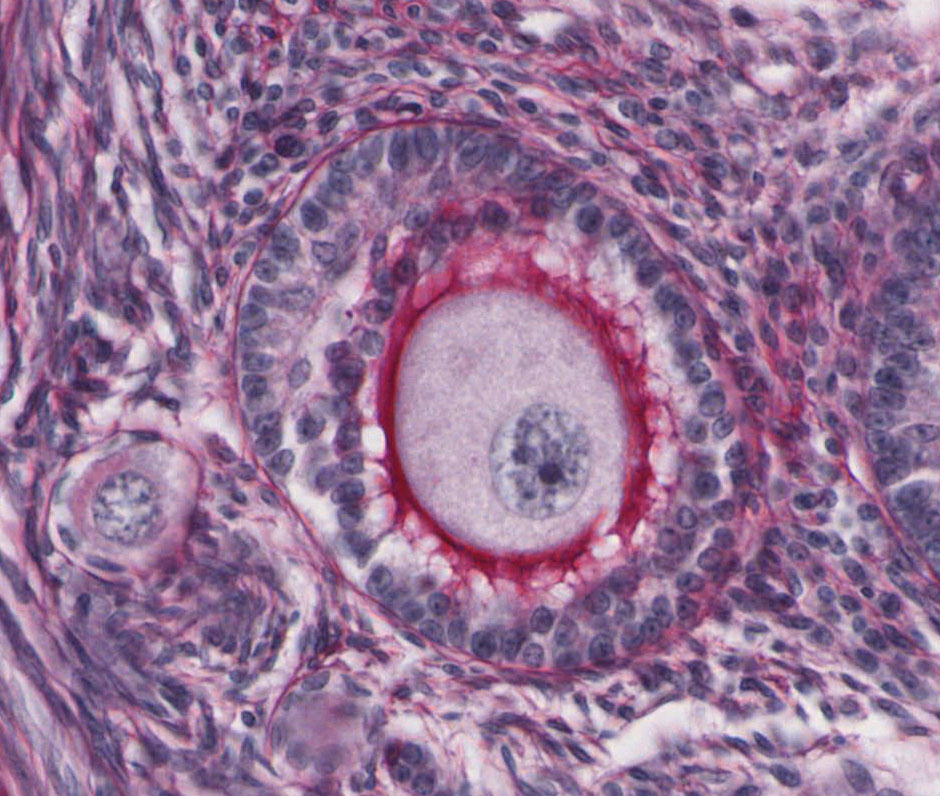
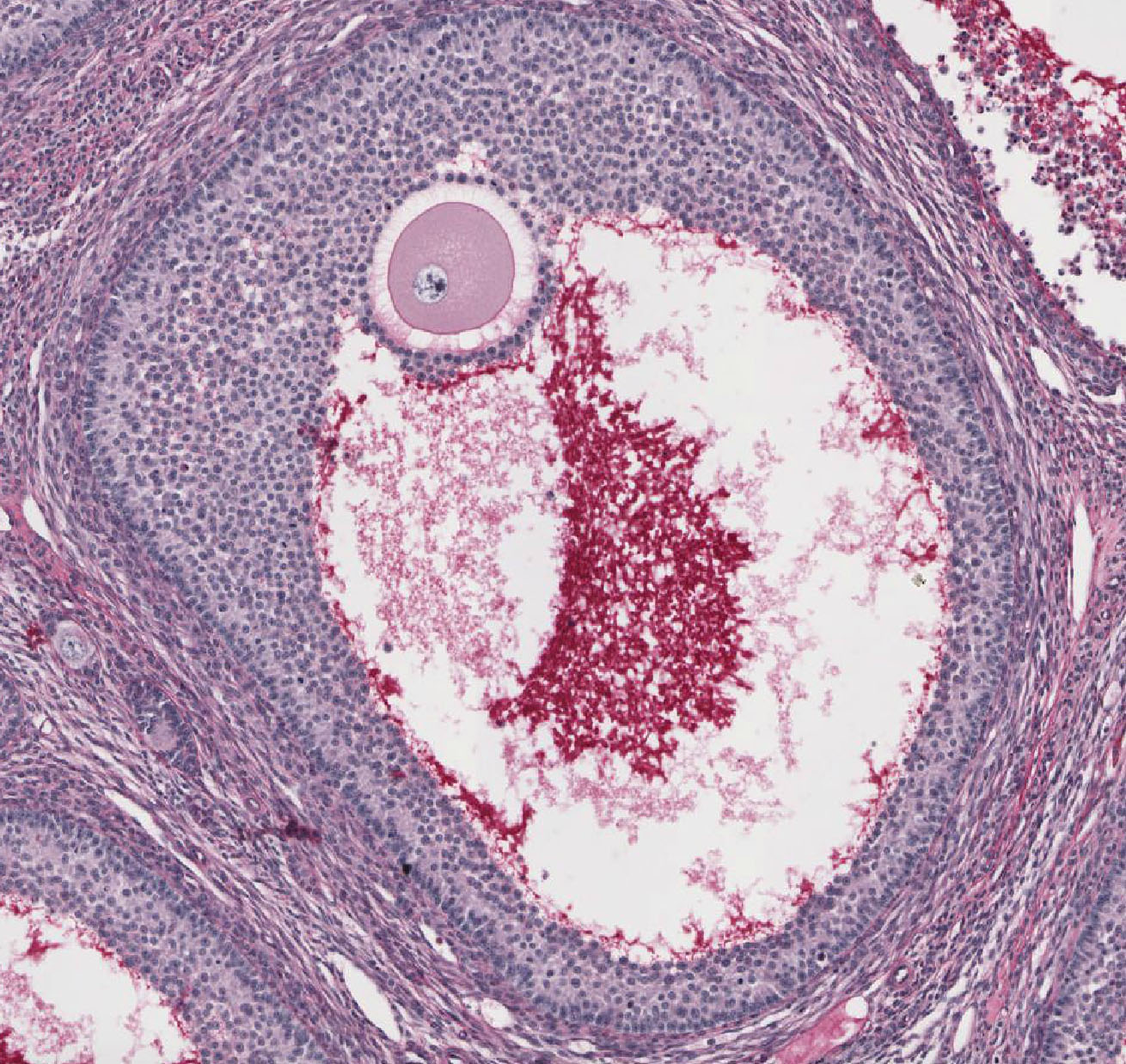
Mature/Graafian Follicle: With continued development, the follicle becomes a Graafian or ovulatory follicle View Image (This follicle is actually rather small to be a real Graafian follicle). The granulosa zone now consists of many layers of cuboidal follicular epithelial cells located at the periphery of the large, well-formed follicular antrum. The oocyte has attained its full size, is located eccentrically within the follicle in a small hillock, the cumulus oophorus which protrudes into the antrum. The zona pellucida is surrounded by a continuous layer of follicular cells, the corona radiata. Because of its size, the oocyte will not be present in every section of the follicle, but examine the other components of a tertiary follicle. The theca interna is separated from the granulosa cells by a distinct basement membrane. Theca externa cells are densely packed, spindle-shaped cells which blend with the theca interna cells and with the surrounding stroma. Note that the theca interna has a rich capillary vascular supply, particularly well demonstrated in slide 235 View Image.
{kind=link}
{kind=link}
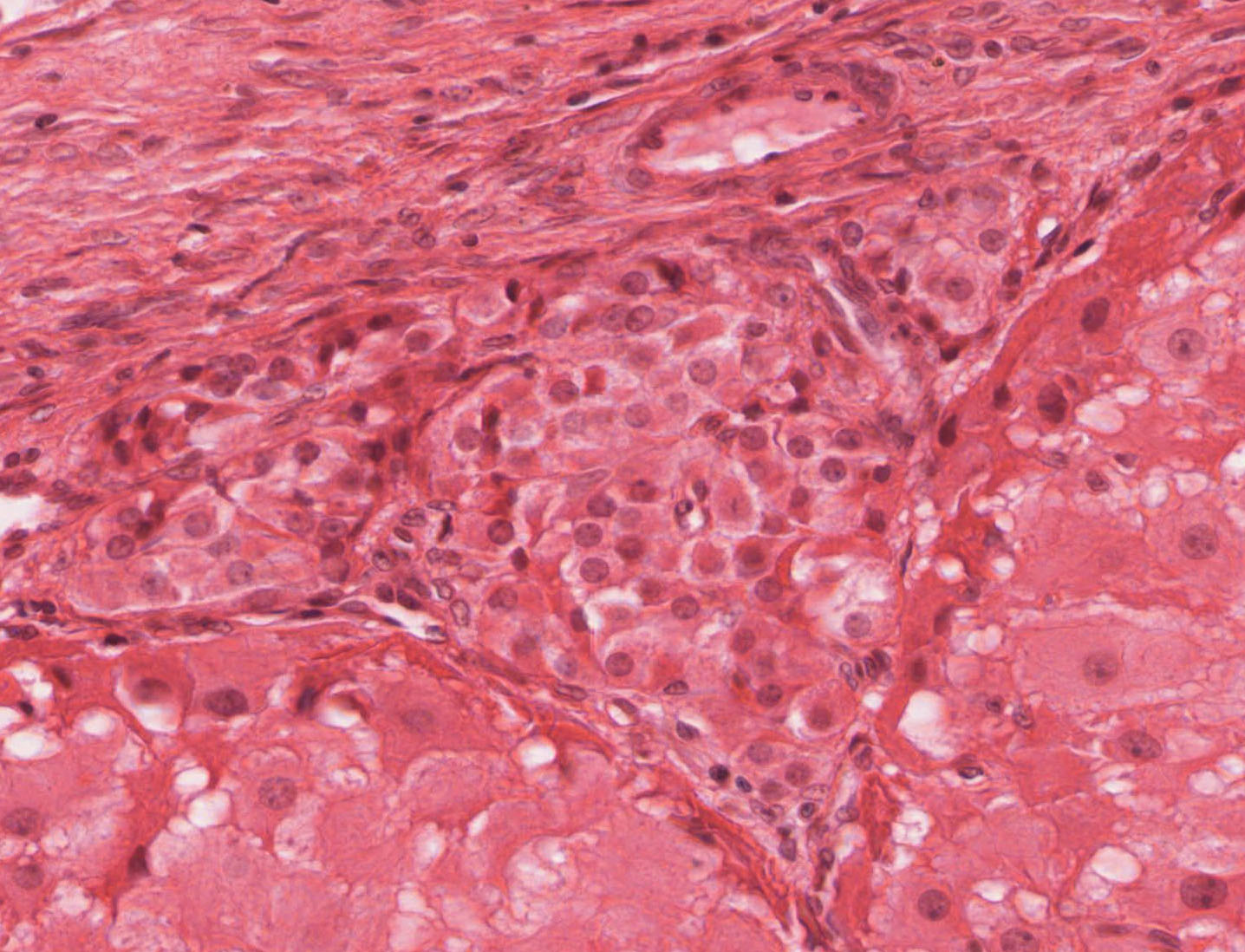
Atretic Follicles: Because the contents of only one follicle are usually ovulated at a time in humans, other follicles which have been stimulated to develop must degenerate, or undergo atresia View Image. Atresia is not limited to mature follicles, but may begin at any stage in follicular development. Early atretic alterations include: clumping of the nuclear chromatin (pyknosis) and shrinkage and lysis of the cytoplasm of the oocyte, granulosa or follicular cells. Examine the pyknotic granulosa cells View Image, which are sloughed into the follicular antrum. The basement membrane that separates the granulosa cells from the theca interna may also thicken considerably to form a so-called “glassy membrane.” These changes are especially well illustrated in H&E slide 234 View Image and trichrome-stained slide #234-2 View Image. Make sure you are able to differentiate atretic changes from artifacts related to shrinkage due to fixation. Macrophages may eventually invade the center of the larger atretic follicles that are finally replaced by loose connective tissue.
{kind=link}
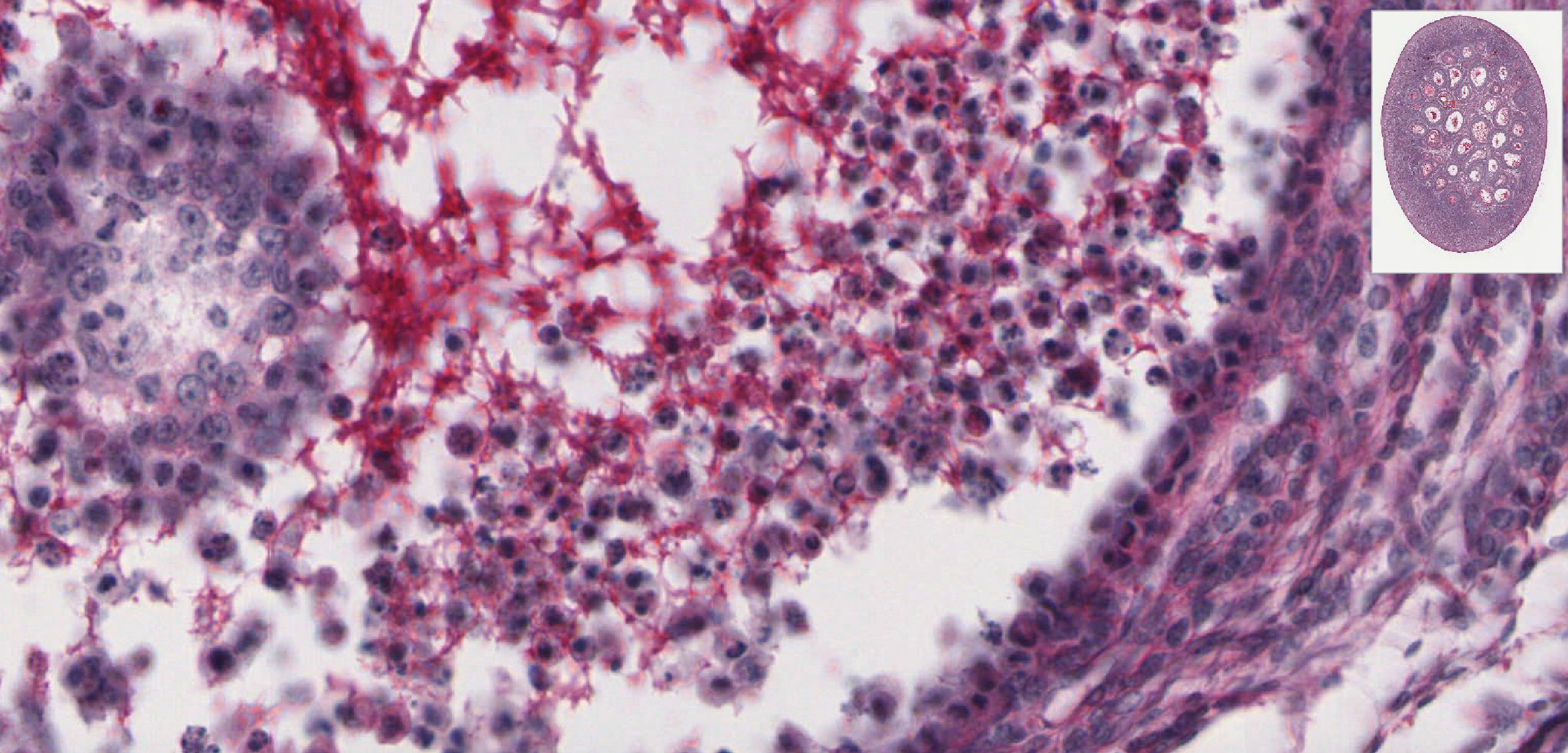{kind=link}
{kind=link}
{kind=link}
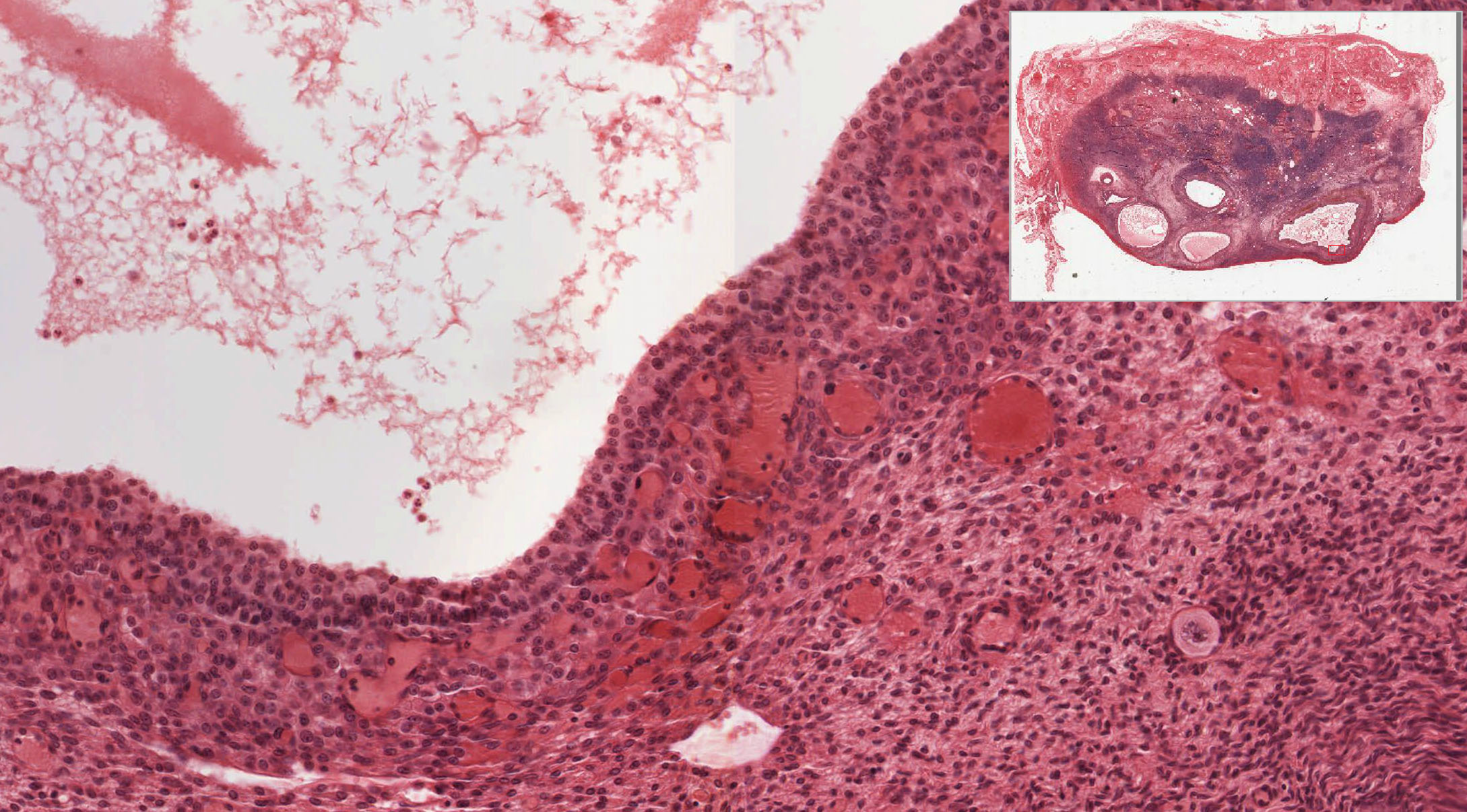
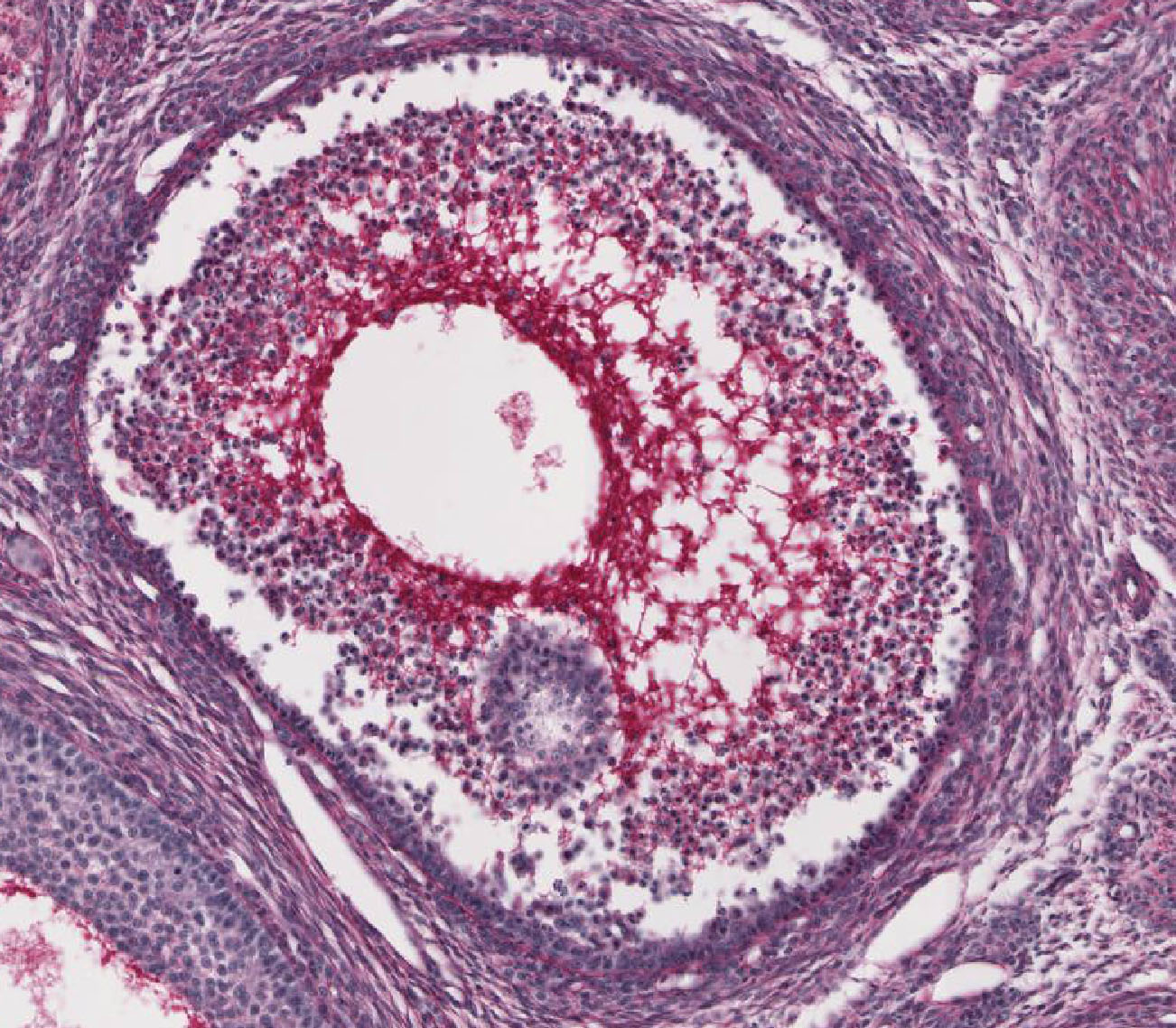
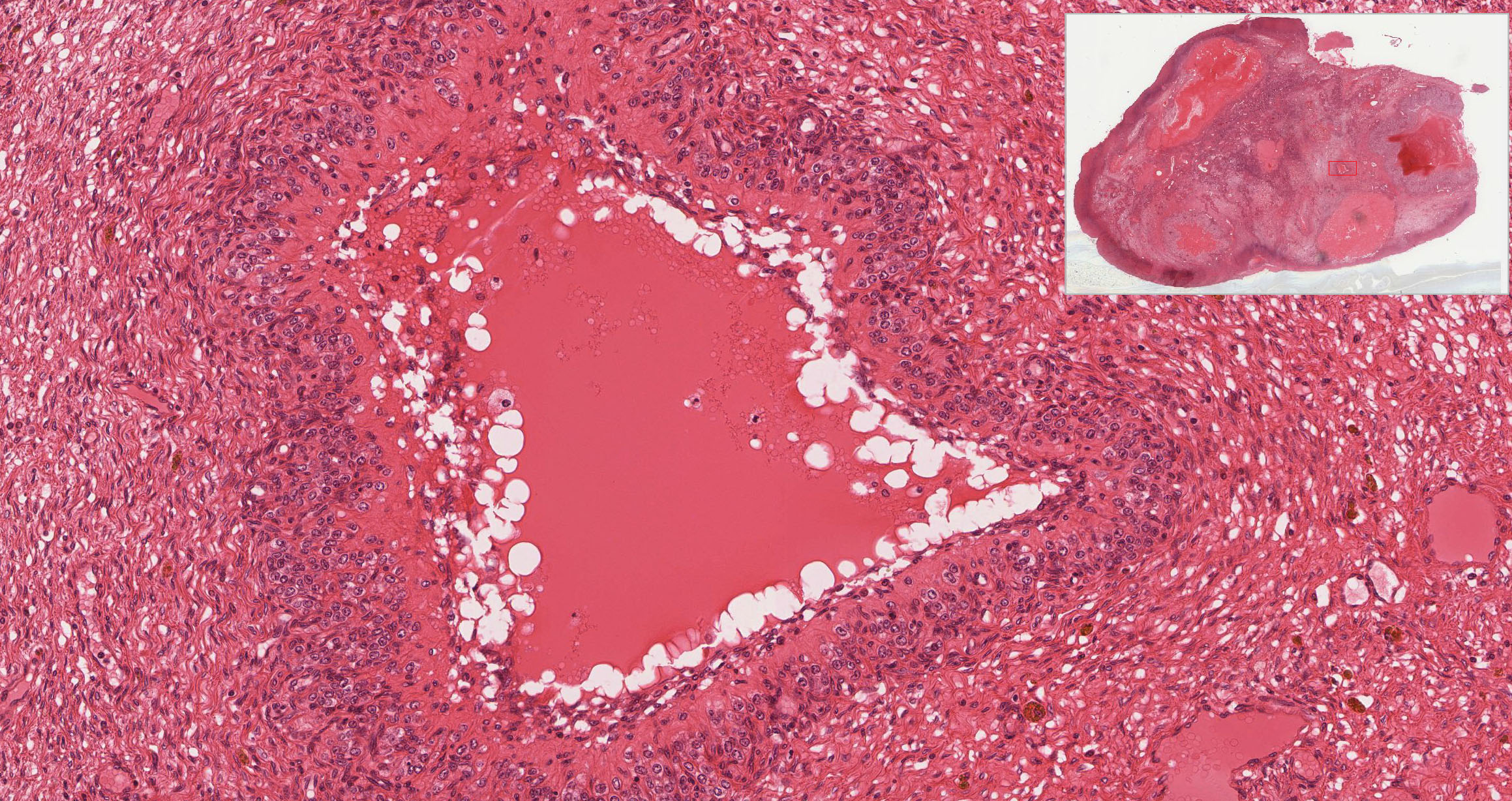
Corpus Luteum: After ovulation, the follicle which housed the ovum collapses and becomes highly infolded and invaded by vessels, forming the corpus luteum View Image (yellow body). Examine slide 236a and observe that the corpus luteum appears pale and very folded. If the egg is fertilized and implants, the corpus luteum enlarges to become the corpus luteum of pregnancy. Examine the inner granulosa lutein cells View Image (formed from the remaining granulosa cells) and the outer theca lutein cells View Image which come from the remaining theca interna cells. Both cell types are polyhedral and filled with lipid droplets and have centrally located nuclei. The theca lutein cells are, however, considerably smaller, more darkly staining and have fewer lipid filled vacuoles than the granulosa lutein cells. They are found most prominently in the infoldings right up against the granulosa lutein layer. Granulosa lutein cells contain a pigment, lipochrome, which produces the yellowish color of the corpus luteum in an unfixed ovary. A central blood clot may be present in recently ovulated follicles.
{kind=link}
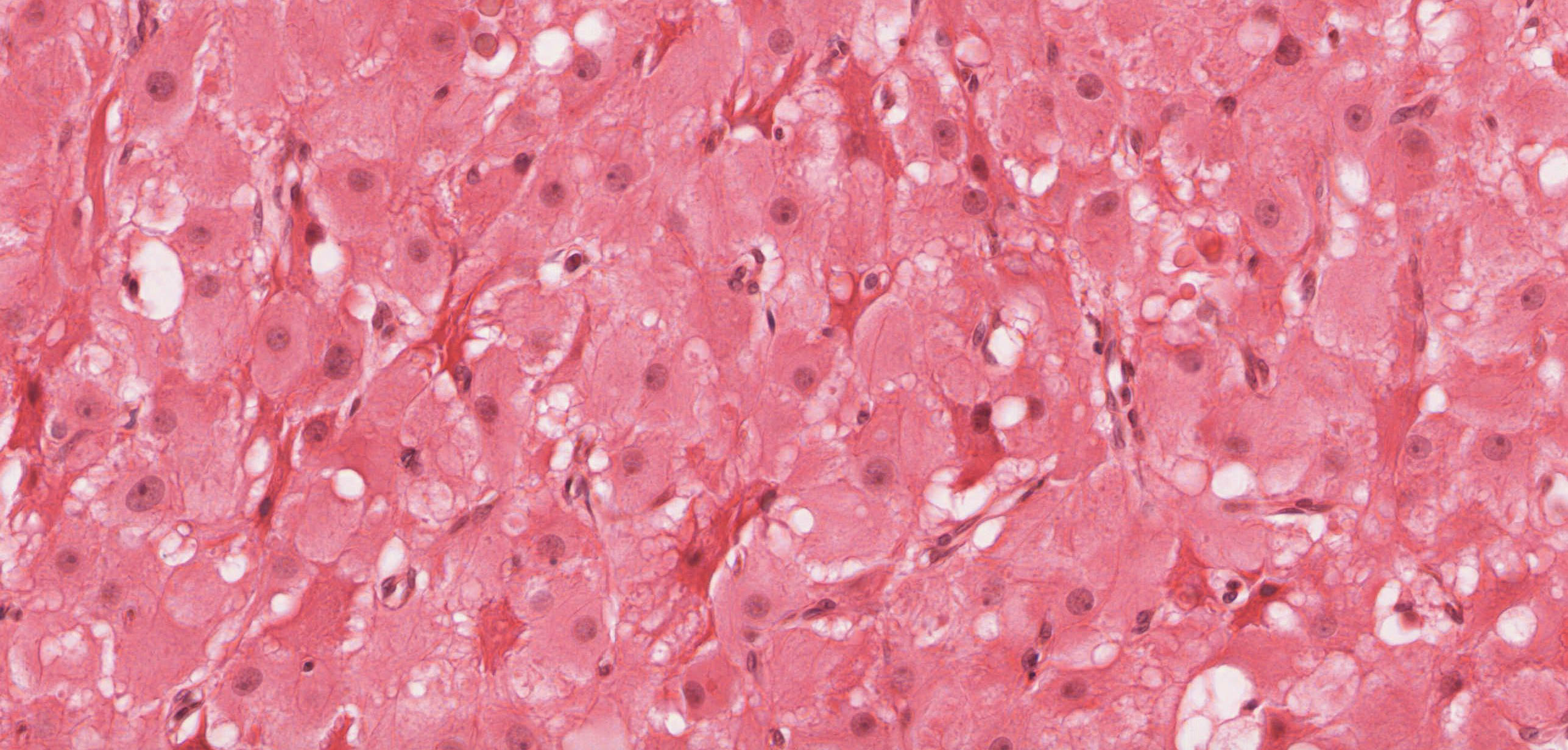{kind=link}
{kind=link}
Corpus Albicans: If the egg is not fertilized, the corpus luteum degenerates, and is gradually infiltrated with collagen and a few (if any) fibroblasts, forming the corpus albicans (white body) particularly evident in slide 234 stained with H&E View Image and trichrome View Image. The corpus albicans is also formed during the later half of pregnancy when the placenta takes over steroid secretion from the corpus luteum. Excellent examples of corpus albicans can be best observed in slides 234 or236.
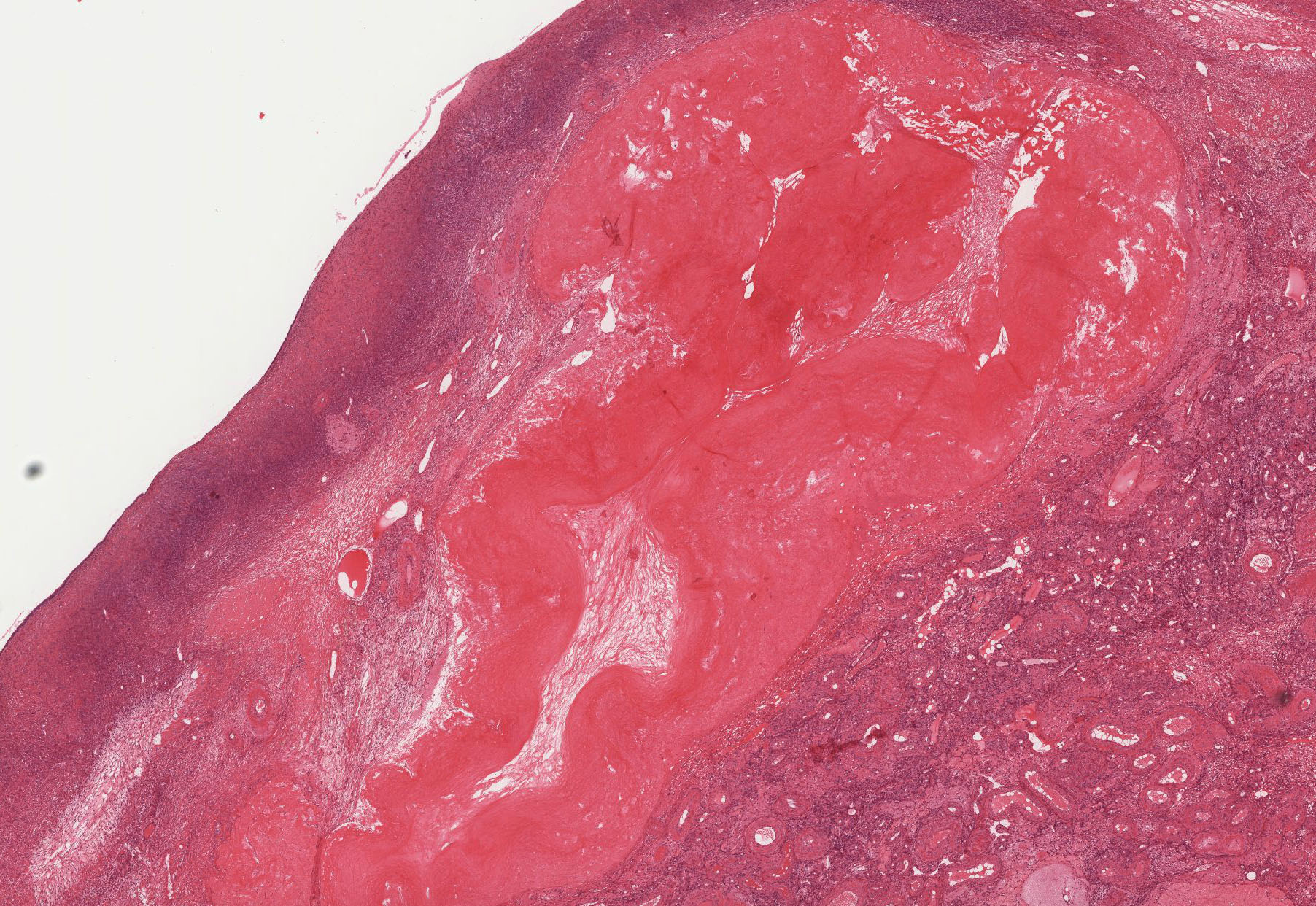{kind=link}
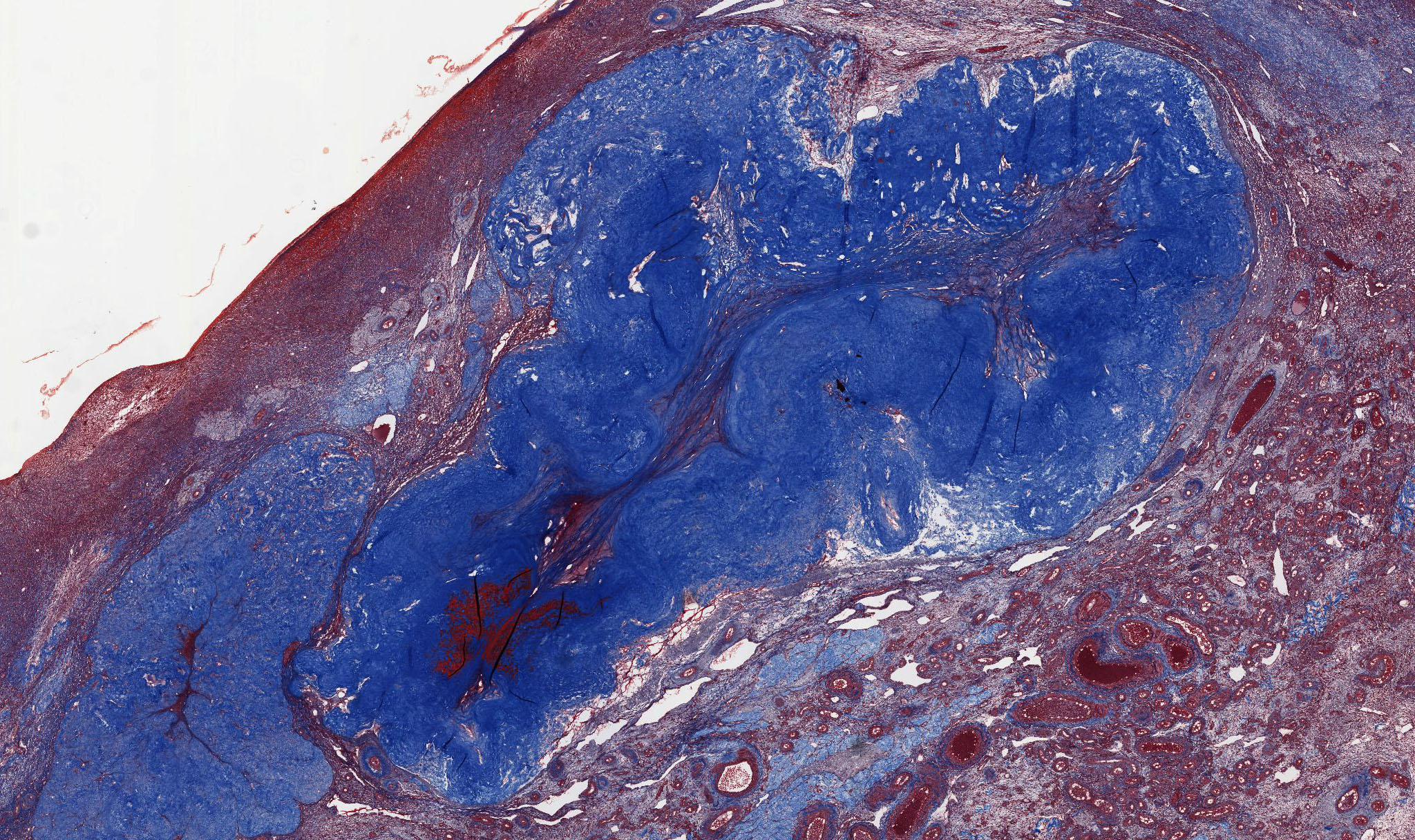{kind=link}
Slide 240-1 Oviduct, infundibulum, H&E View Virtual Slide
Slide 240-2 Oviduct, ampulla, H&E View Virtual Slide
Slide 241even Oviduct, isthmus, H&E View Virtual Slide
Slide 241odd Oviduct, uterine segment, H&E View Virtual Slide
The oviduct conducts the ovulated egg from the peritoneal cavity to the uterus over a period of approximately three days, during which fertilization and segmentation of the zygote occurs. There are striking regional variations in the oviducts that reflect:
1) the trapping of the egg, 2) a region where fertilization takes place, and 3) a region simply for transport to the uterus.
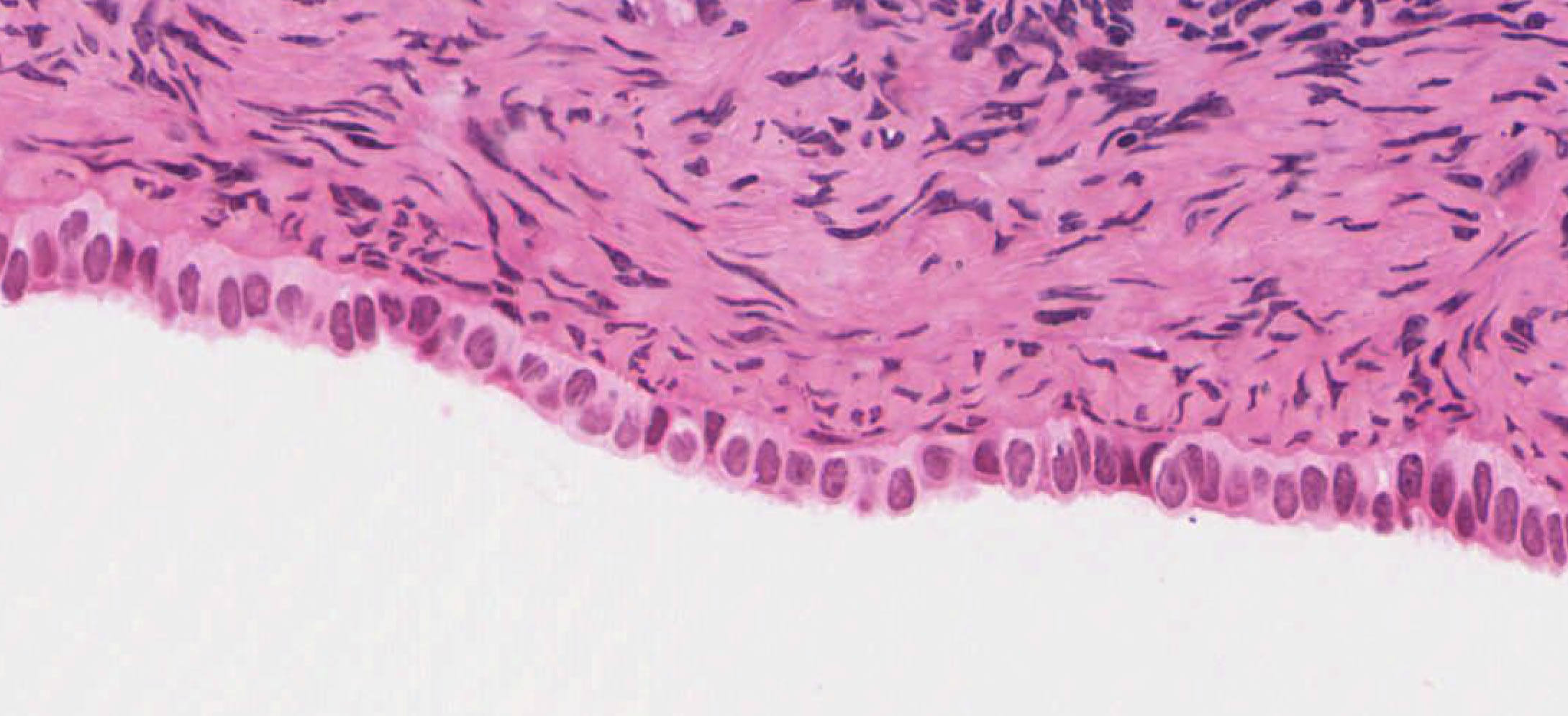
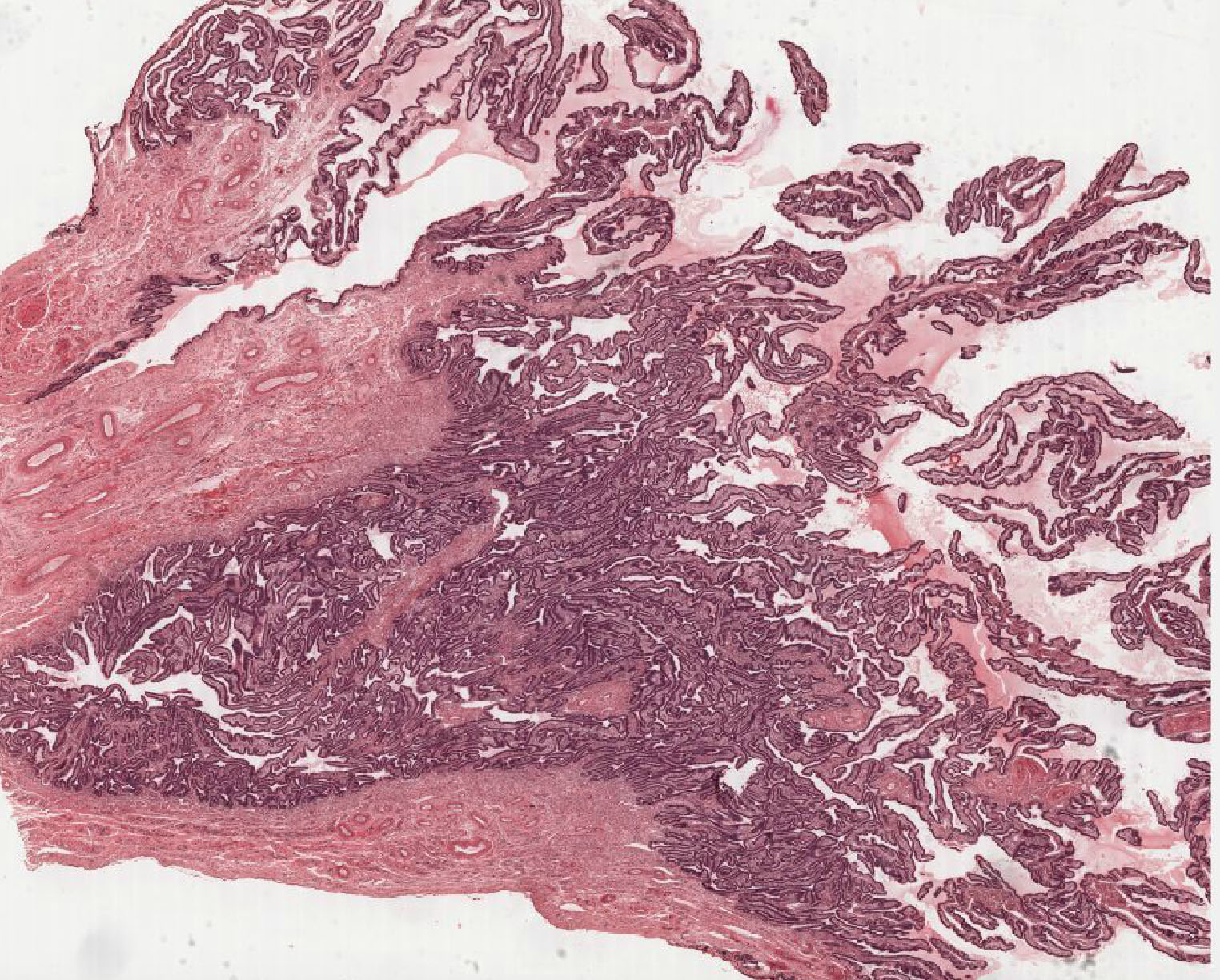
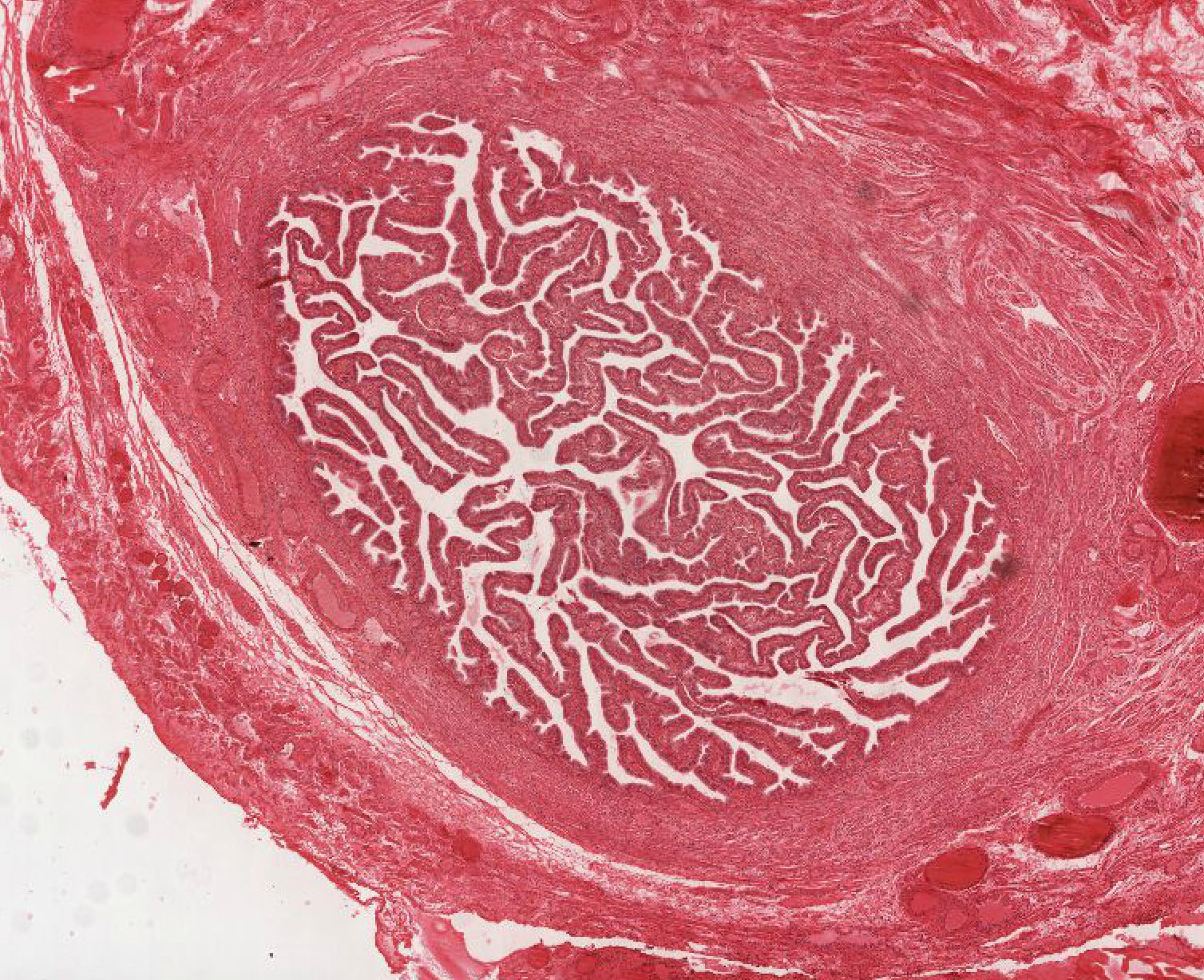
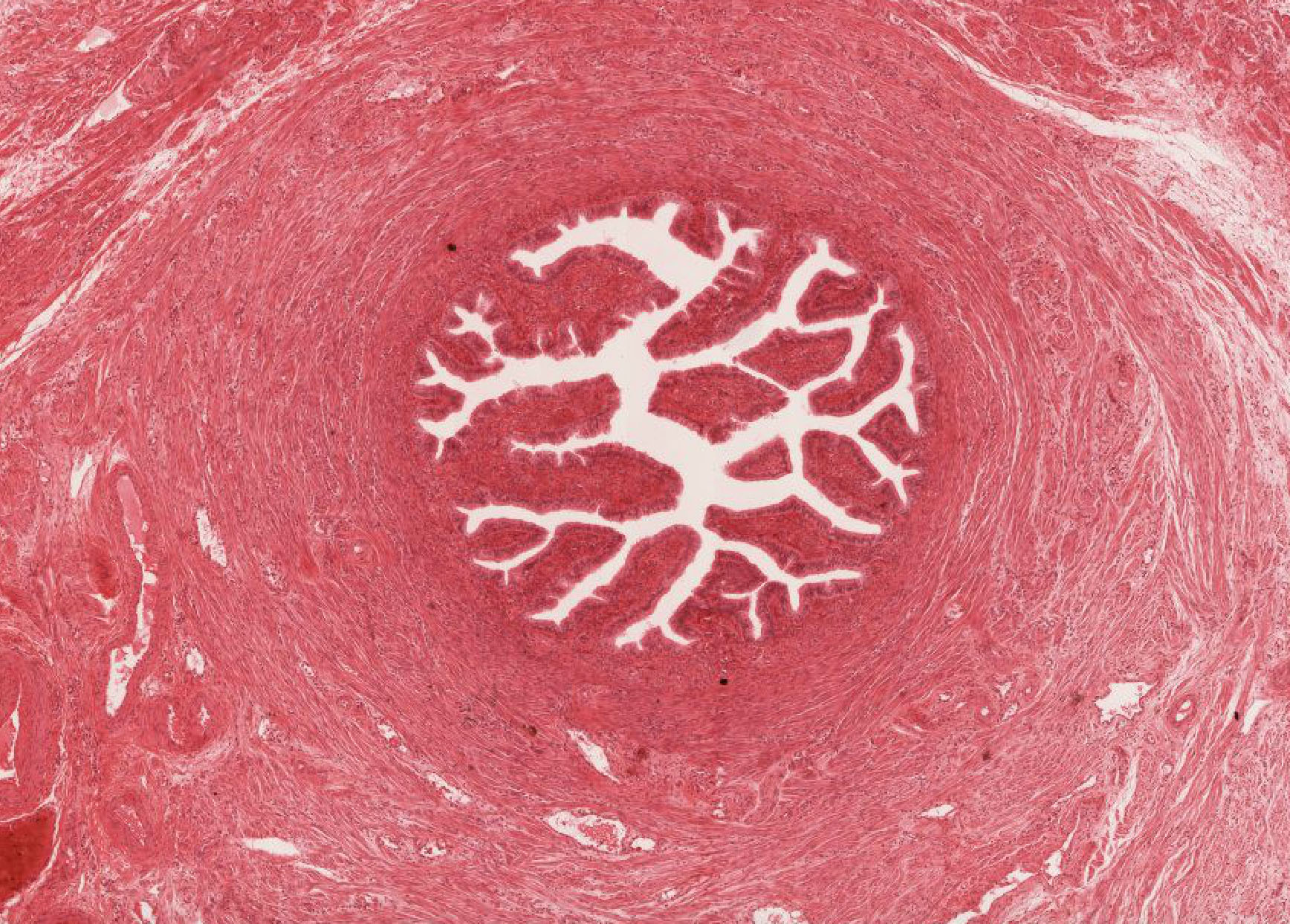
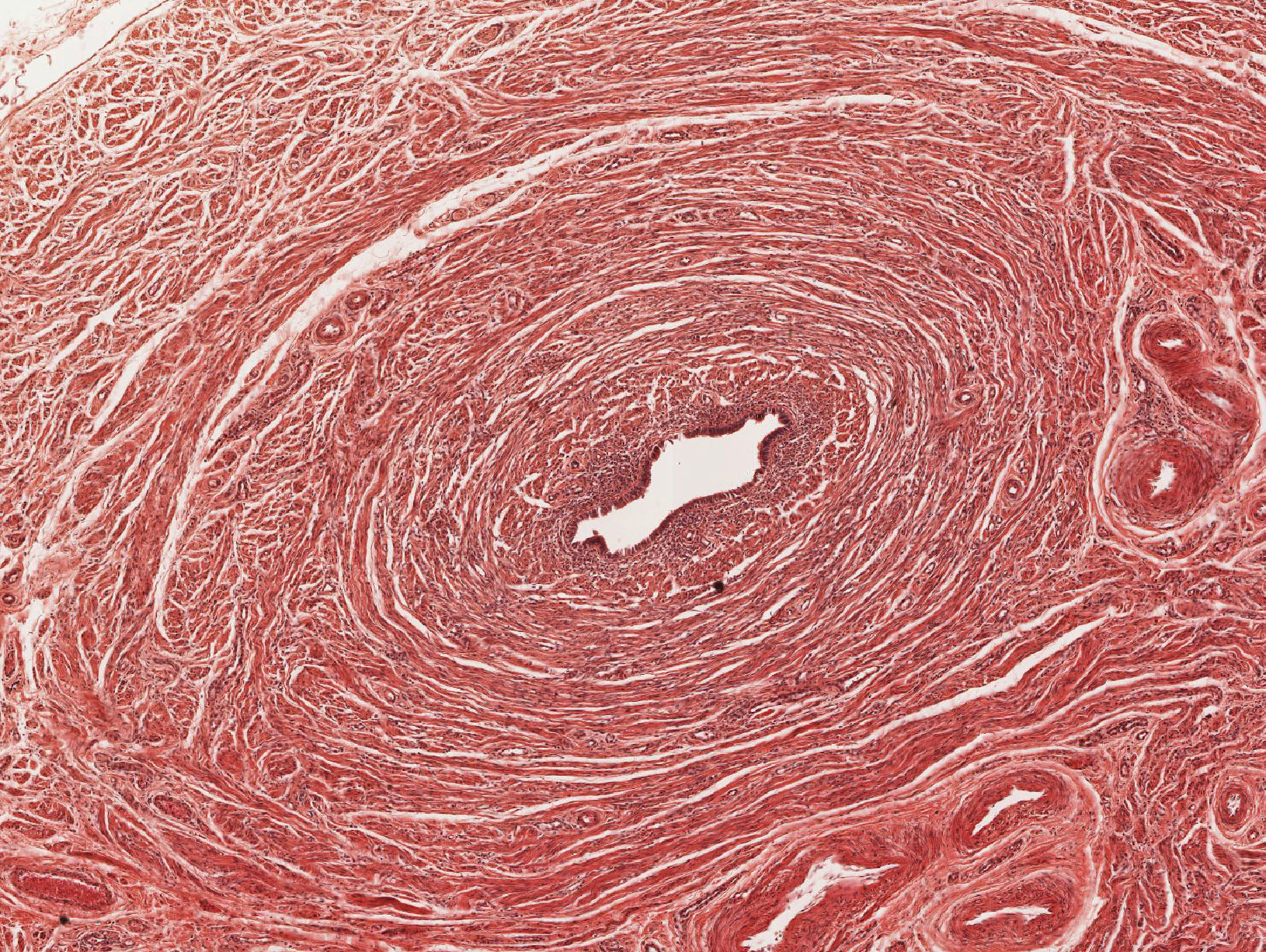
Examine its open end near the ovary, the infundibulum View Image, in slide 240-1 and note the funnel shape and ridges of mucosa that extend as a fringe of finger-like projections known as fimbriae. Note the ciliated cells that cover the fimbria that help sweep the egg into the oviduct. Move on to slide 240-2 to trace the passage of the egg from the infundibulum through the numerous longitudinal folds of the ampulla View Image recalling that this is where fertilization usually occurs. Next is the isthmus region in slide 241even View Image that is characterized by tall branching ridges of mucosa which project into and partially fill the lumen of the oviduct. Note the numerous non-ciliated secretory (peg) cells and the ciliated cells that line the lumen. The isthmus contains tall folds and few ciliated cells, while theinterstitial (or intramural because it's within the uterine wall) segment, shown in slide 241odd, View Image has low ridges and a preponderance of secretory cells. Observe that as you progress from the infundibulum where there is a large lumen filled with many mucosal folds toward the uterus, the overall luminal diameter decreases, as does the number of ciliated cells, while the proportion of smooth muscle in the muscular layer gradually increases.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Examine the loose connective tissue of the lamina propria using slide 241even. Next, study the muscularis organized roughly into circular, oblique, and sparse longitudinal layers of smooth muscle. It has been suggested that this muscle may play a two-fold role; first, in distending the oviduct toward the ovary to aid in “capturing” the ovum; and second, in producing peristaltic waves which help propel the ovum toward the uterus. The outermost layer of the oviduct, the serosa, is highly vascular and is composed of mesothelium and thin underlying connective tissue that is continuous with the broad ligament.
279 Ovary - Primordial Follicle View Virtual EM Slide
Note an incomplete layer of flat granulose cells around the oocyte.
280 Ovary - Primary Oocyte View Virtual EM Slide
Note the highly organized appearance of the primary follicle and the considerable size difference to a primordial follicle.
281 Ovary - Secondary Follicle View Virtual EM Slide
In this low magnification view of a secondary follicle, examine the characteristics of the thecal layer, granulosa cells, corona radiata and the ooplasm. Note particularly the basal lamina which separates the thecal and granulosa cell layers and the acellular zona pellucida that surrounds the oocyte.
282 Ovary - Zona Pellucida of a Secondary Follicle View Virtual EM Slide
Higher magnification view of another secondary follicle. Note the penetration of the zona pellucida by processes from the oocyte and from the granulosa cells of the corona radiata. One current theory regarding follicular atresia suggests that contact of the ooplasm by granulosa cells is required for continued follicular development and that, if lost, atresia results.
283 Ovary - Theca Interna and Externa of a Secondary Follicle View Virtual EM Slide
The cells of the theca interna will eventually differentiate into endocrine cells and show characteristics of steroid-producing cells. The theca externa consists of a mixture of smooth muscle cells and connective tissue.
285 Ovary - Corpus Luteum View Virtual EM Slide
You do not need to be able to identify theca and granulose lutein cells based on their EM appearances. However, note the difference in their cytoplasmic contents and know their secretory products.
288 Oviduct Epithelium of the Oviduct View Virtual EM Slide
Cilia and their basal bodies are clearly visible in the apical cytoplasm of the ciliated cells. Note short microvilli on the non-ciliated secretory cells.
Click on a question to reveal the answer.
What is the purpose of the corona radiata?
The corona radiata consists of granulosa cells that have gap junctions with the ovum and therefore likely provide metabolic support. When the ovum is ovulated, the corona radiata and a few additional layers of adhering granulosa cells (collectively called the "cumulus mass") remain adhered to the ovum, probably continuing to support the ovum and adding bulk to allow for transport of the ovum down the oviduct via peristalsis and the beating action of ciliated cells lining the oviduct.
What is the morphological difference between the corpus albicans and an atretic follicle?
The corpus albicans is formed after the corpus luteum stops producing hormones and a new cycle begins. It is a fibrous mass that is broken down by macrophages in the ovary. Histologically it is very large and a predominant feature of the ovary. Follicular atresia occurs in all of the developing follicles that do not become the dominant follicle. It involves a thickening of the basement membrane between the theca interna and granulosa cells. Because of the smooth appearance of its thickened basement membrane, atretic follicles are also called the glassy membrane. Atretic follicles are generally much smaller than the corpus albicans.
What is the correlation between the follicular stage and the ovarian stage during the menstrual cycle?
In females, the process of gamete production begins well before birth. Within the first month as an embryo, primordial germ cells enter the gonad and begin dividing by mitosis. As birth approaches, these germ cells enter prophase of meiotic division and arrest in that stage. These are known as primary oocytes. The fetus can have upwards of seven million primary oocytes. Primary oocytes have the normal number of chromosomes, but they have twice normal amounts of DNA (here n refers to number of chromosomes and c refers to the amount of DNA in a single set of chromosomes). Before birth and up until puberty, the primary oocytes are contained in primordial follicles, which have a single layer of flat cells surrounding them and are located in the cortex of the ovary.
At puberty, both lutenizing hormone (LH) and follicle stimulating hormone (FSH) begin to affect the ovum. FSH induces follicular growth and maturation from a primordial follicle to a primary follicle with a thicker layer of surrounding cells. The oocyte is still a primary oocyte with double normal DNA (2n, 4c).
FSH continues to stimulate the follicle to become a secondary and then mature or Graafian follicle. At this stage, right before ovulation, the ovum completes its first meiotic division and becomes a 1n, 2c cell (called a secondary oocyte). Though the genetic material is divided evenly between the two cells, the cytoplasm goes almost completely to one of the cells. It is this cell that continues on. The other is known as a polar body (in this case, the first polar body).
After ovulation, the follicle becomes the corpus luteum. The secondary oocyte begins its second meiotic division at this point and it stops in the metaphase stage. It continues to be a 1n, 2c cell.
If the ovum remains unfertilized, nothing more happens. If, however, fertilization occurs, then the ovum progresses. After the sperm penetrates the egg, the ovum completes its second meiotic division and a second polar body is formed. It is at this point that the egg becomes a 1n, 1c cell and is prepared to combine with the genetic material of the sperm. The table below summarizes each stage (and the corresponding ploidy) of oocyte development.
- Distinguish the cyclical alterations in the uterine endometrium and understand their hormonal bases.
- Describe the alterations and functional changes in the cytology of the cervix and vagina during the menstrual cycle, and during pregnancy.
Slide 244 Uterus, proliferative phase, H&E View Virtual Slide
Slide 243 Uterus, early secretory phase, H&E View Virtual Slide
Slide 245-1 Uterus, secretory phase, H&E View Virtual Slide
Slide 245-2 Uterus, menstrual phase, H&E View Virtual Slide
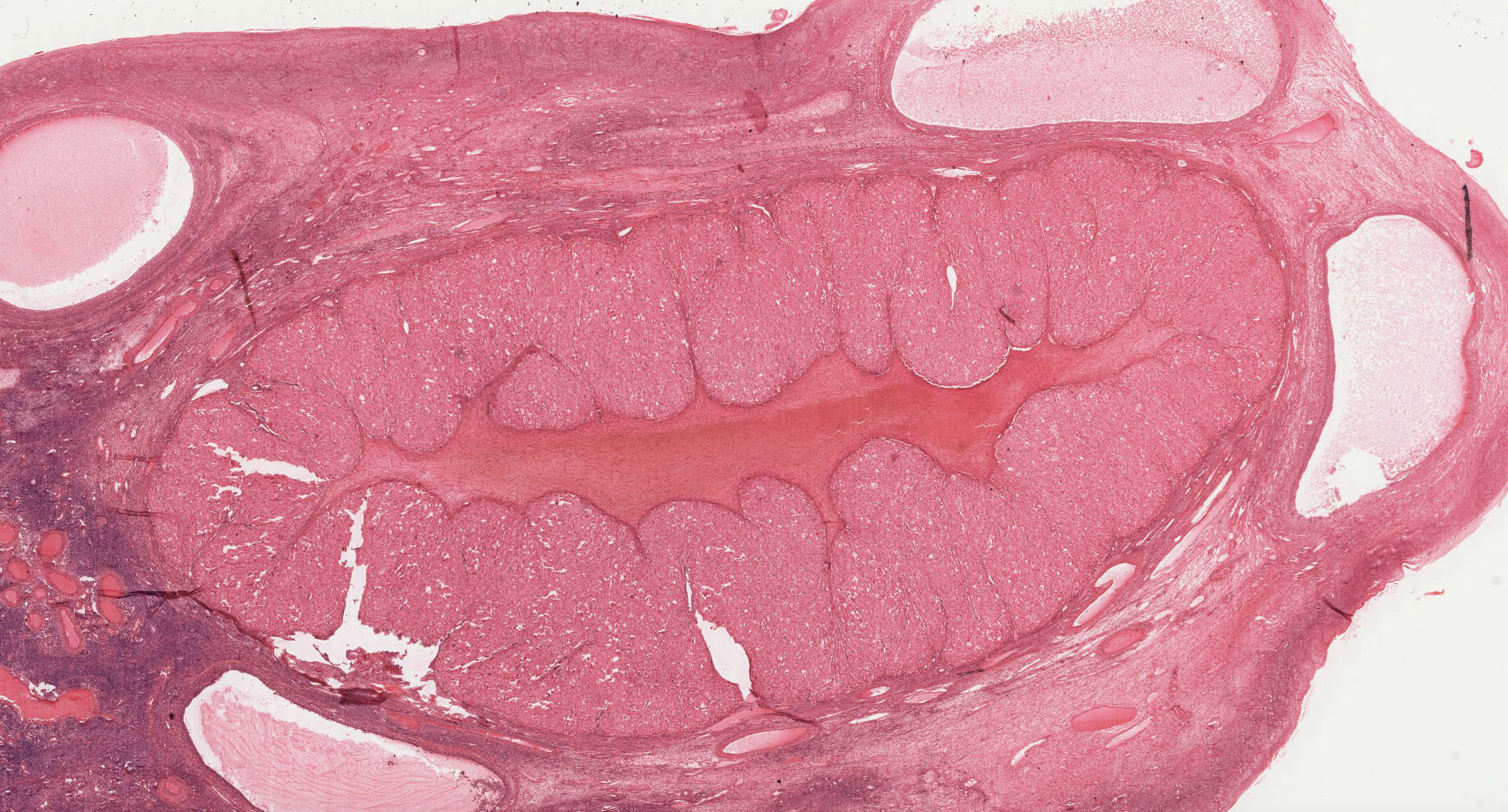
The uterus is a pear-shaped, hollow organ partially covered by peritoneum. Examine the overall topographical organization of the uterus from its outer serosal or adventitial covering, through the thick muscular layer (myometrium) to the mucosa (endometrium) in slide 243 (this slide is actually at a stage between proliferative and secretory).
The myometrium View Image of the uterus is composed of interwoven bundles of smooth muscle supported by dense connective tissue. Examine their arrangement into indistinct bundles, with several bundles running roughly parallel to the long axis of the body of the uterus. During pregnancy, the muscle fibers elongate, proliferate and produce collagen in response to hormonal stimulation.
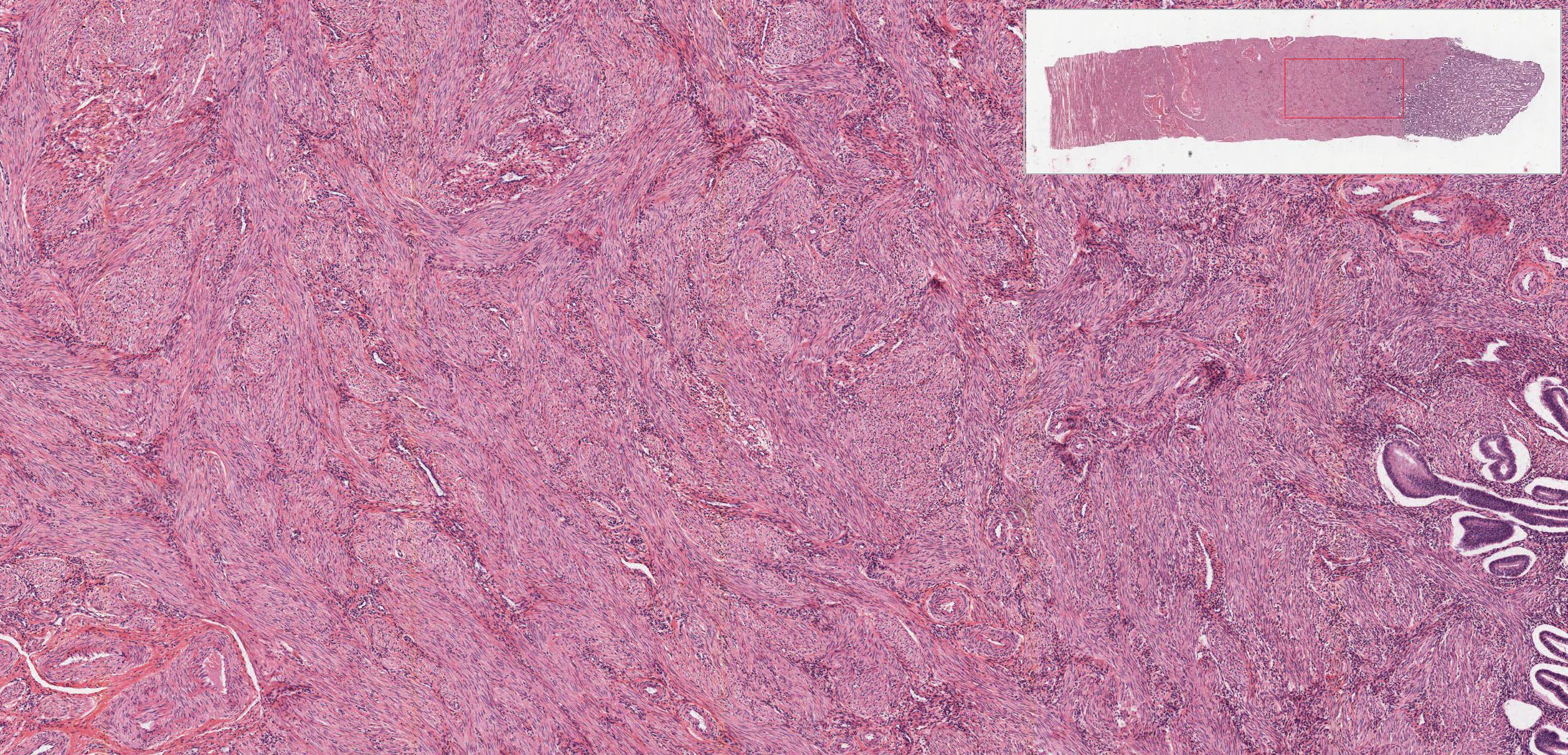{kind=link}
The endometrium View Image is divided into two layers or strata and consists of a simple columnar epithelium and an underlying stroma that contains tubular glands that may branch near the myometrium. The stratum functionale or functional layer is the thicker, outer layer that is sloughed off with menstruation. The underlying stratum basale or basal layer is retained throughout the menstrual cycle and serves as the regenerative source of the stratum functionale. The endometrium undergoes striking cyclical modifications that depend on sequential hormonal alterations. Compare specimens from proliferative and secretory phases with the menstrual phase.
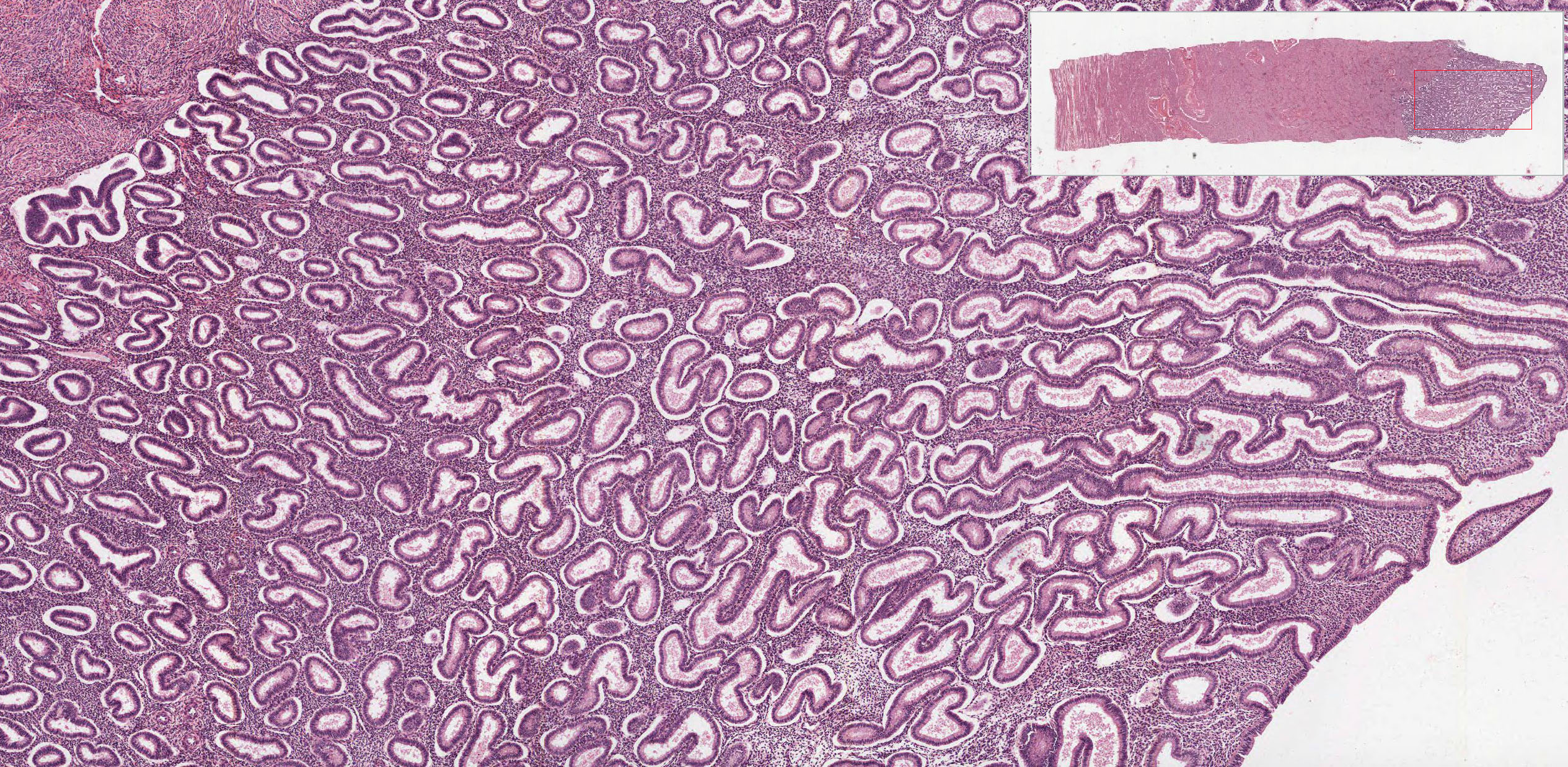{kind=link}
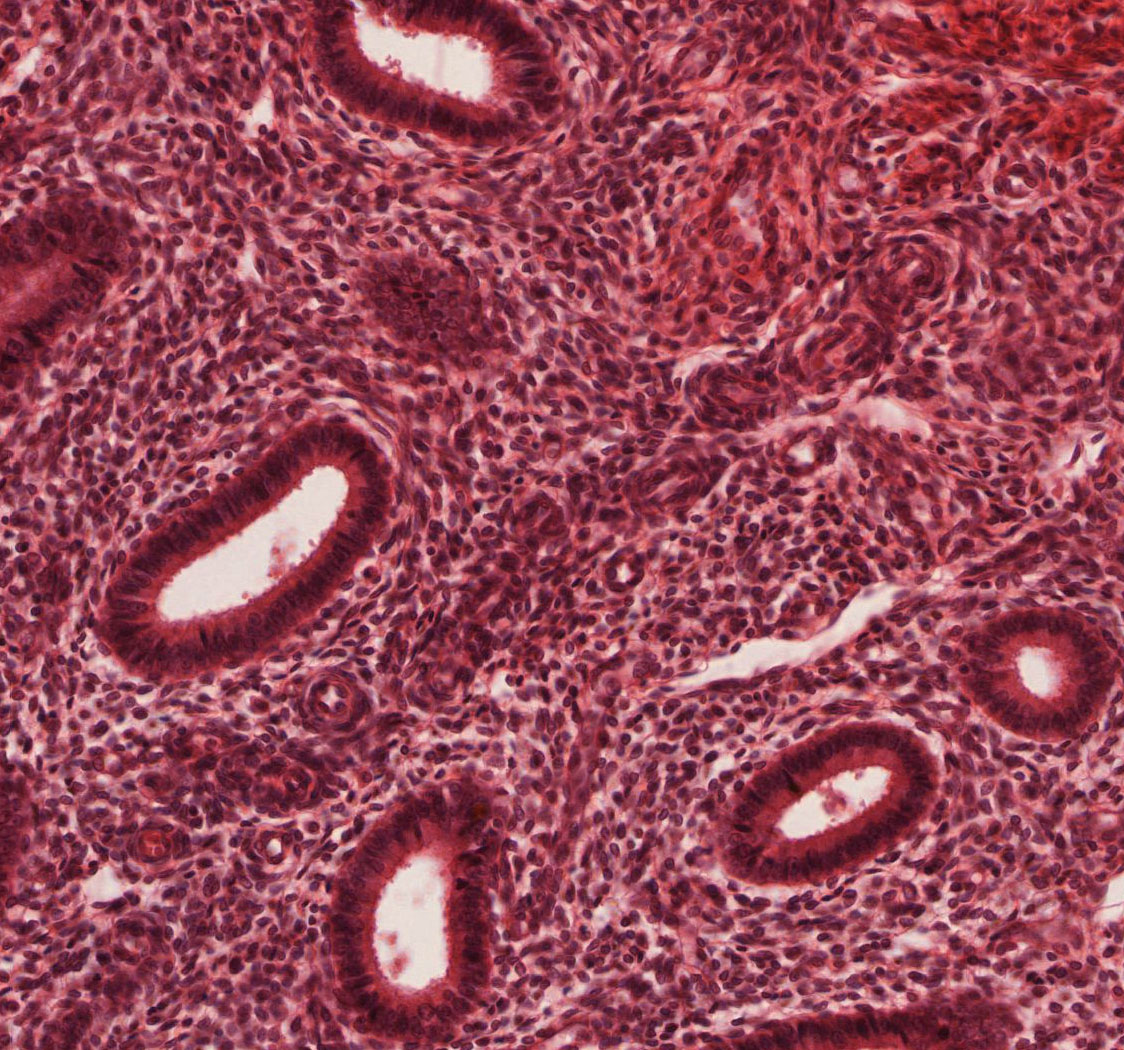
- Proliferative Phase (days 5-14; W pg 370, 19.15; pg 372, 19.18) In slide 244 View Virtual Slide, the proliferative phase is underway and glands have proliferated and cover the surface. Although difficult to see in these sections, spiral arteries View Image are elongated and convoluted, and extend from the basal layer into the functional layer.
- Secretory Phase (days 15-28; W pg 373, 19.19) In slide 245-1 View Virtual Slide, observe that the glands of the secretory phase endometrium appear convoluted. Toward the end of this phase, apical tissues become ischemic and glands take on a characteristic “saw tooth” appearance View Image. The endometrium reaches its maximal thickness during this period, and spiral arteries continue to grow and extend into the superficial regions of the functional layer. There is also considerable leukocyte infiltration in the stroma.
- Menstrual Phase (days 1-4; W pg 371, 19.16) In slide 244-2 View Virtual Slide, much of the stratum functionalis has sloughed away, and amongst the debris are numerous blood cells (RBCs and leukocytes). Notice that the stratum basalis remains intact.
{kind=link}
{kind=link}
Slide 249 Cervix, H&E View Virtual Slide
UCSF slide 405 Cervix, trichrome stain View Virtual Slide (virtual slide courtesy of the University of California, San Francisco)
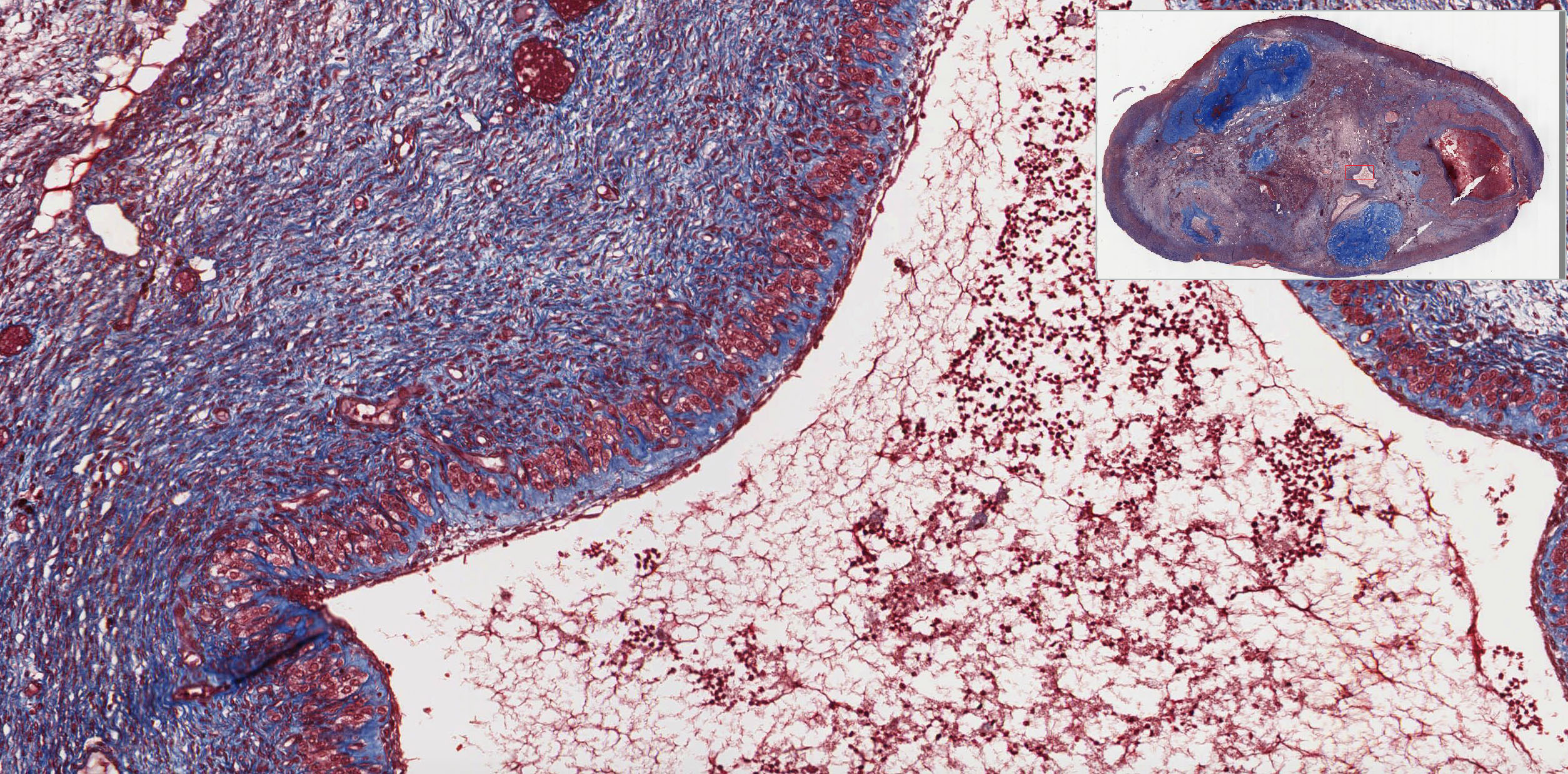
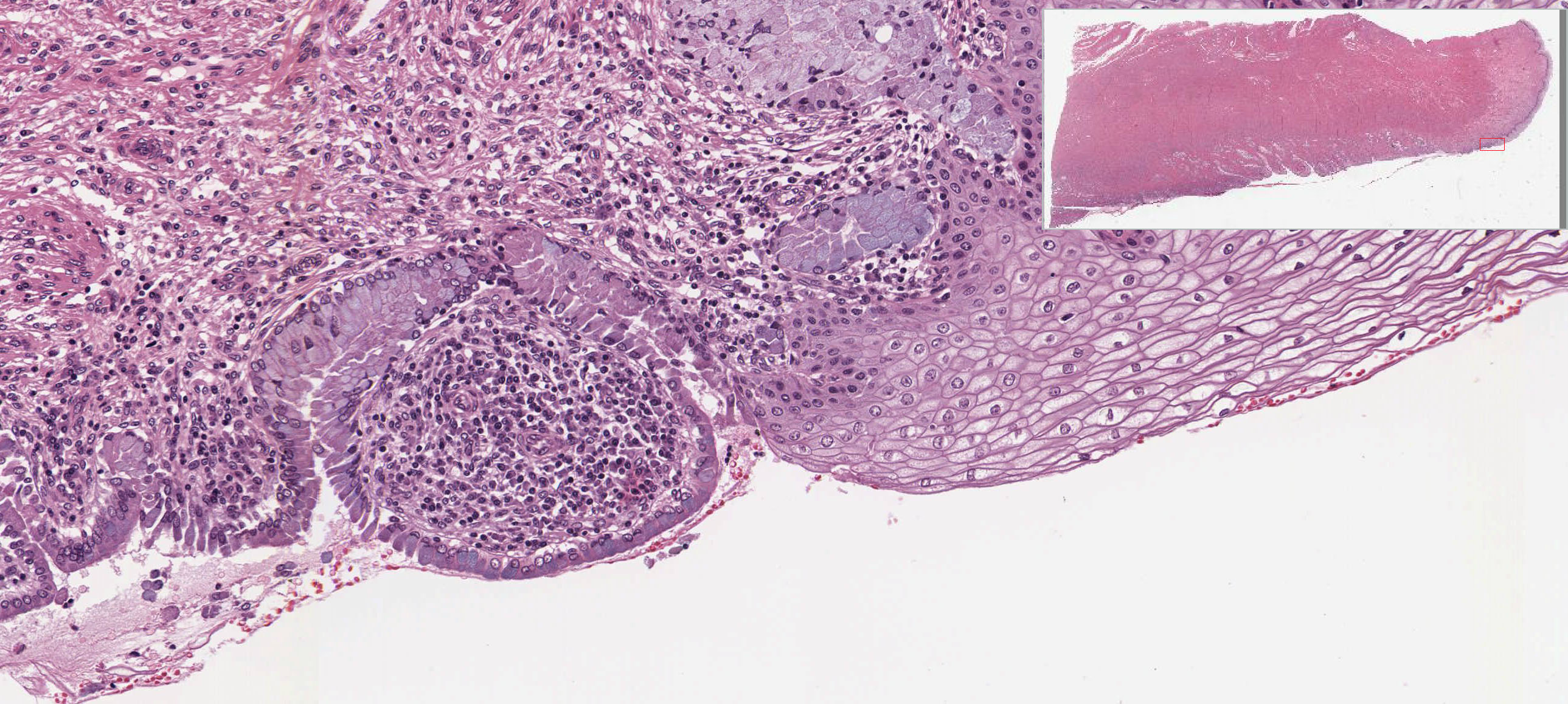
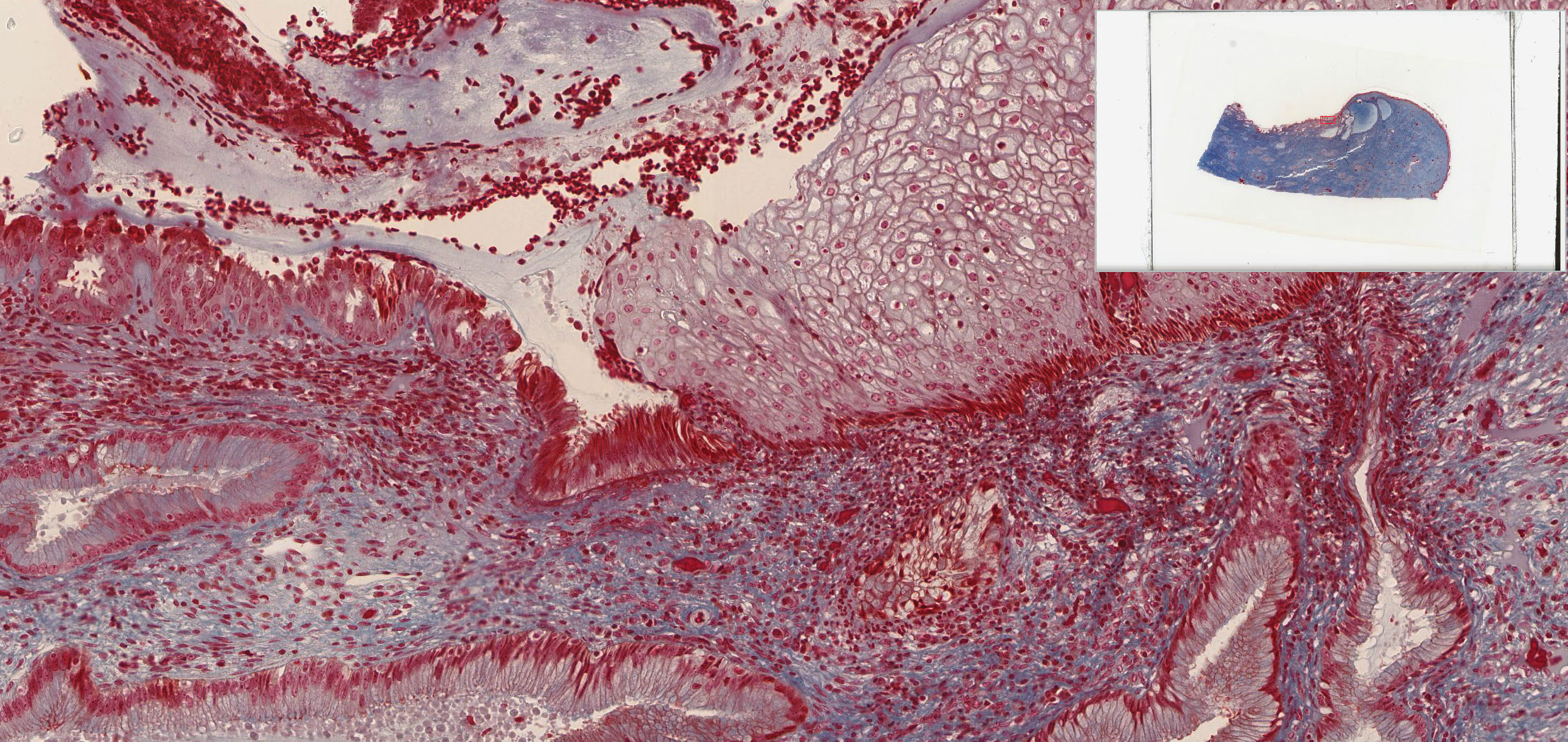
The uterine cervix shown in slide 249 is continuous with both the body of the uterus and the upper portion of the vagina. Note that the wall has considerable smooth muscle and much dense connective tissue. Note also the number of collagen fibers in the stroma.
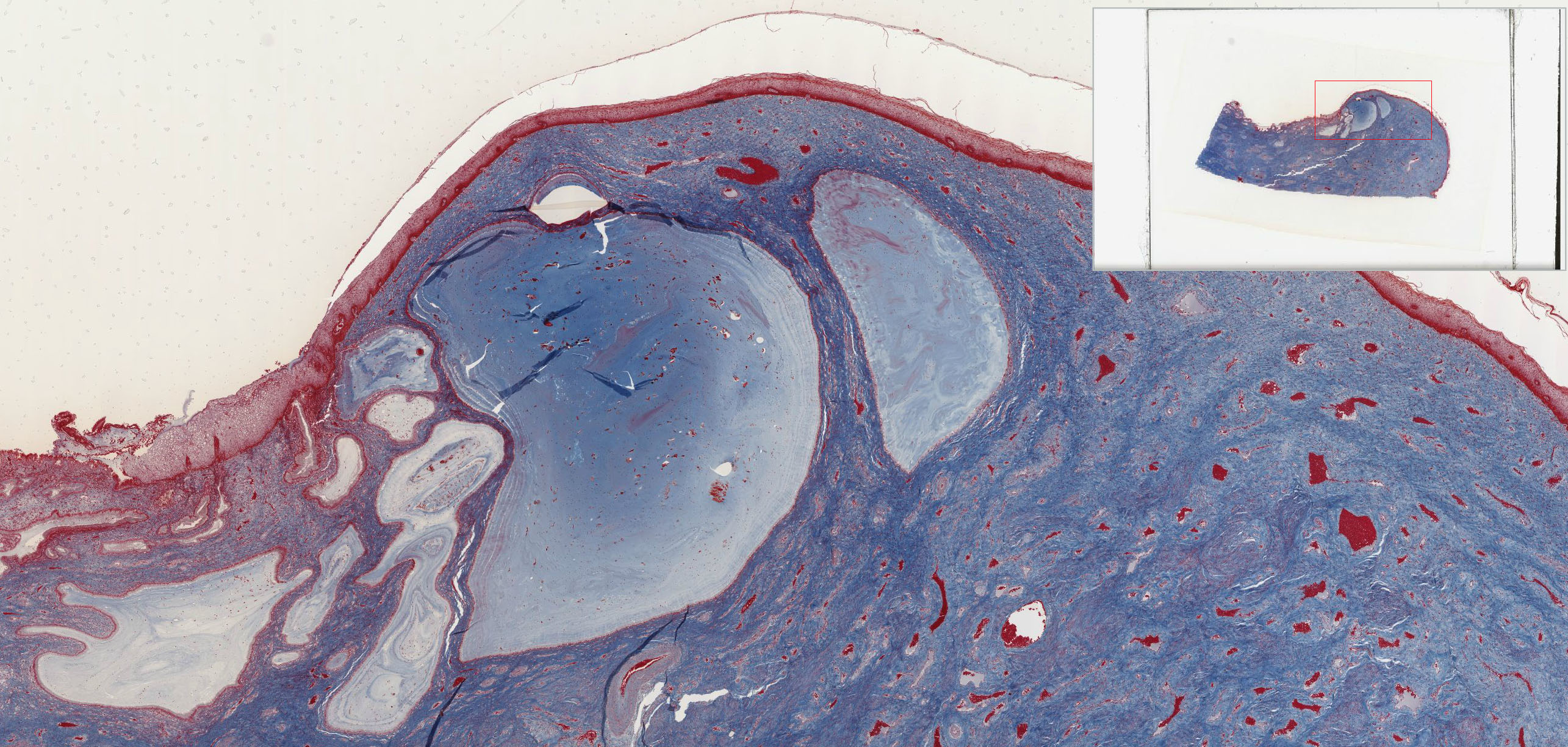
The mucosa is lined by a tall columnar mucus-secreting epithelium in its uterine portion, but note the abrupt change to stratified squamous epithelium at its vaginal face. This stratocolumnar junction which should be readily identifiable in both slide 249 View Image and UCSF slide 405 View Image is frequently the site of pre-neoplastic and neoplastic (cervical cancer) changes. The mucosa is thrown into deep irregular folds known as plicae palmitae (palmate folds). During the majority of the uterine cycle these glands secrete a highly viscous mucus forming a barrier to microorganisms, while at mid-cycle (ovulation) the mucus becomes more hydrated, which facilitates sperm entry. Blockage of the openings of the cervical mucosal glands frequently results in the accumulation of secretory products within the glands, leading to the formation of dilated Nabothian cysts which may be seen in USCF slide 304 View Image. These cysts are generally benign; however, they can become clinically relevant should they become enlarged enough to cause obstruction of the cervical canal.
{kind=link}
{kind=link}
{kind=link}
Slide 250-1 Vagina, H&E View Virtual Slide
Slide 250-2 Vagina, trichrome stain View Virtual Slide
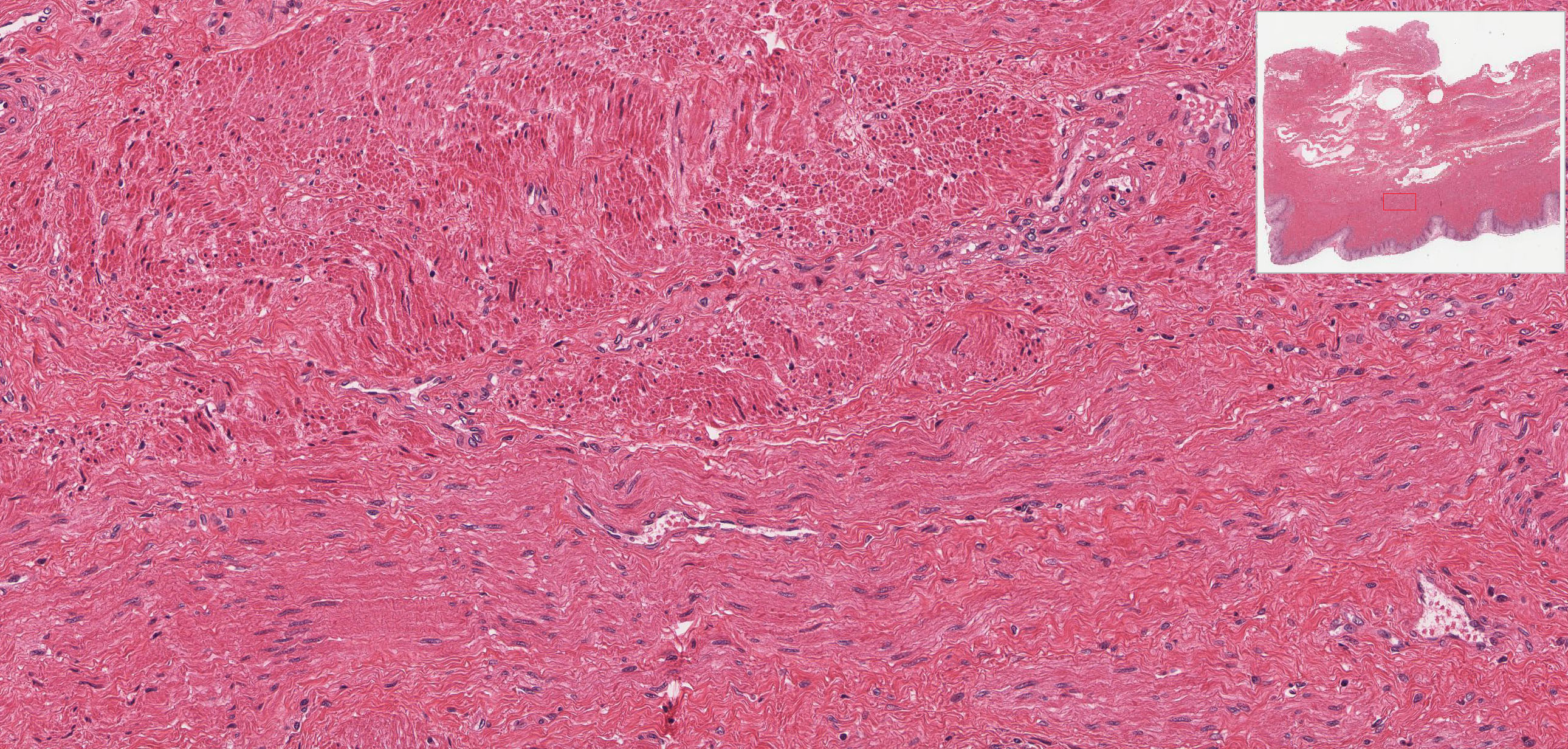
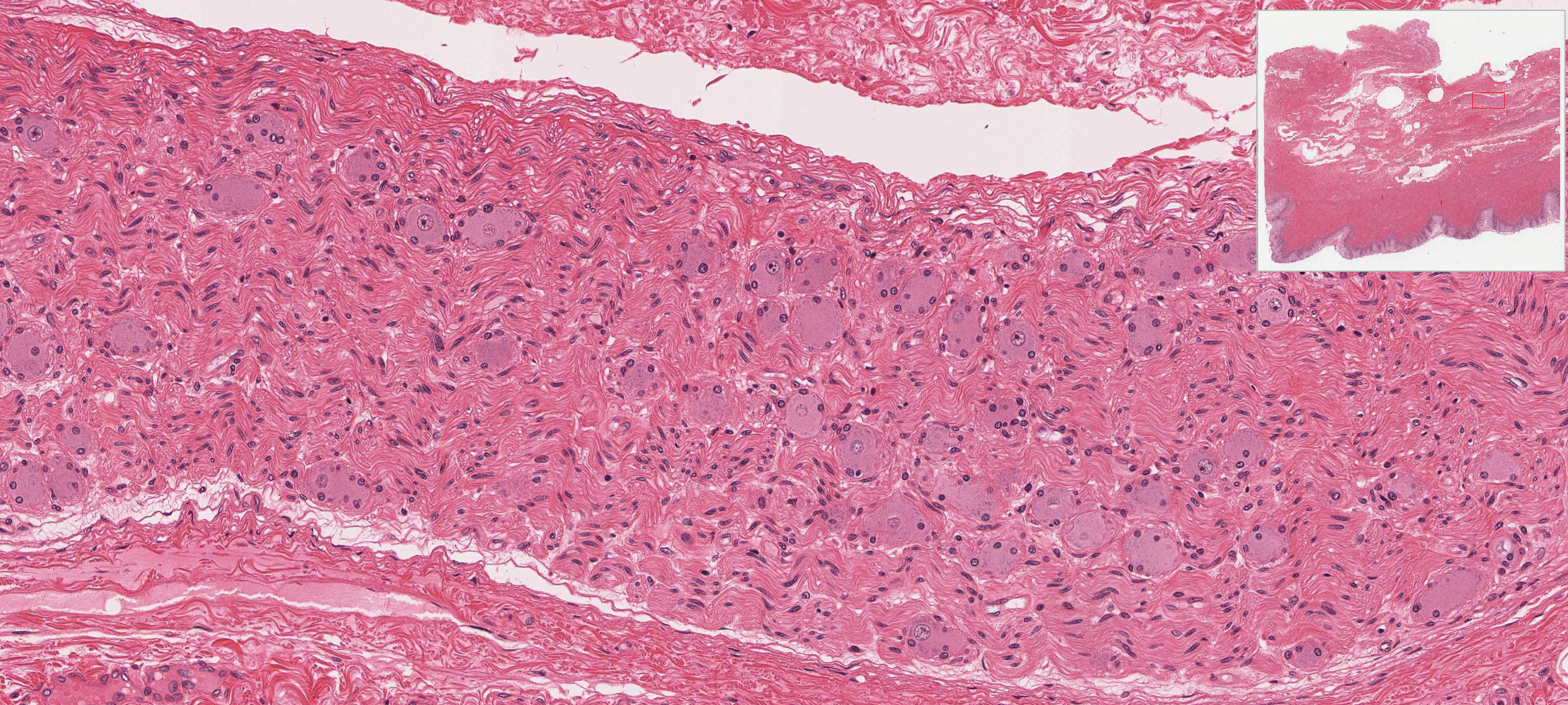
The vagina connects the female reproductive system to the exterior of the body. At low magnification, note its three-layered structure: the mucosal lining, smooth muscle layer, and outer adventitial layer. The epithelium that lines the vagina is stratified squamous, which appears slightly vacuolated due to the presence of glycogen in these cells, which is removed during fixation.
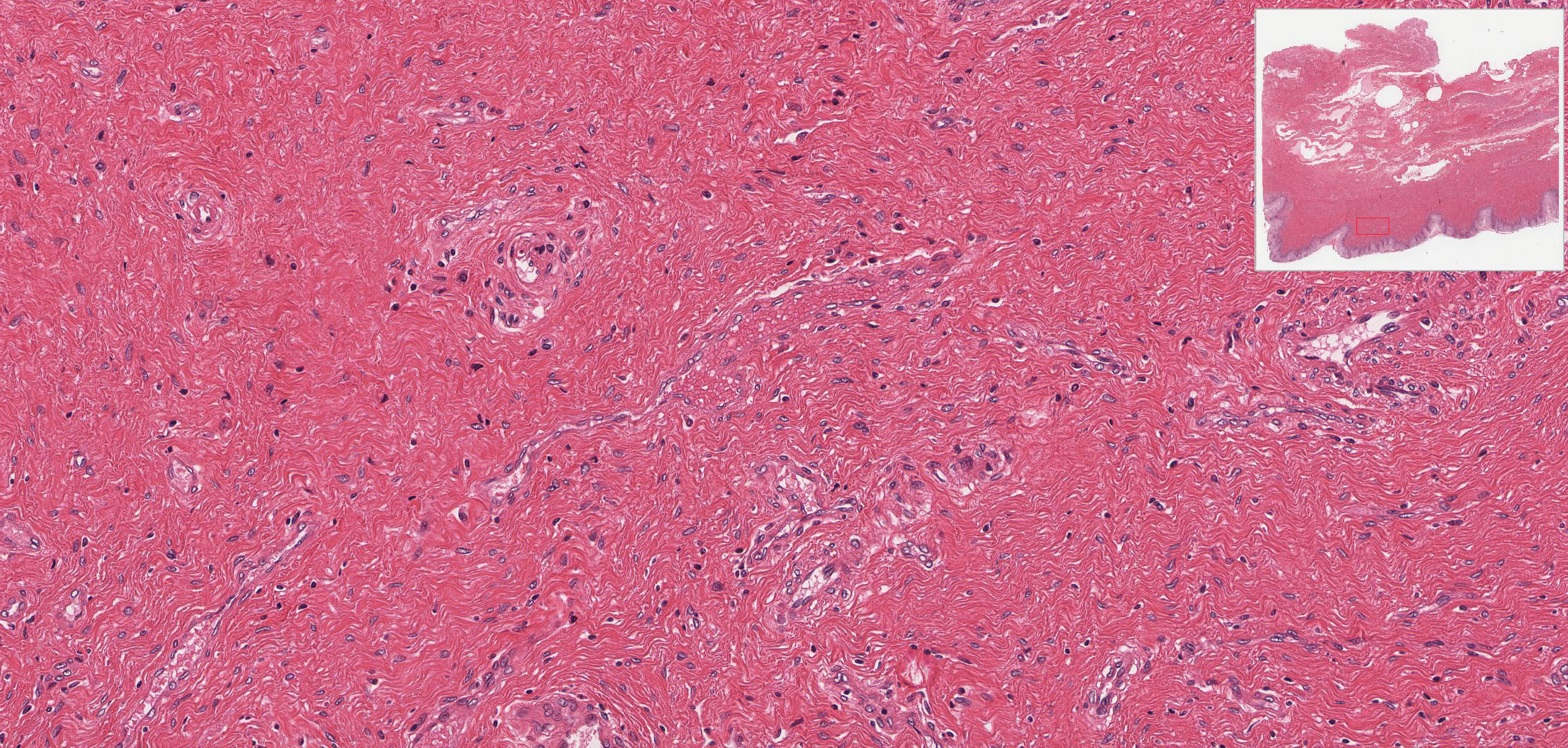
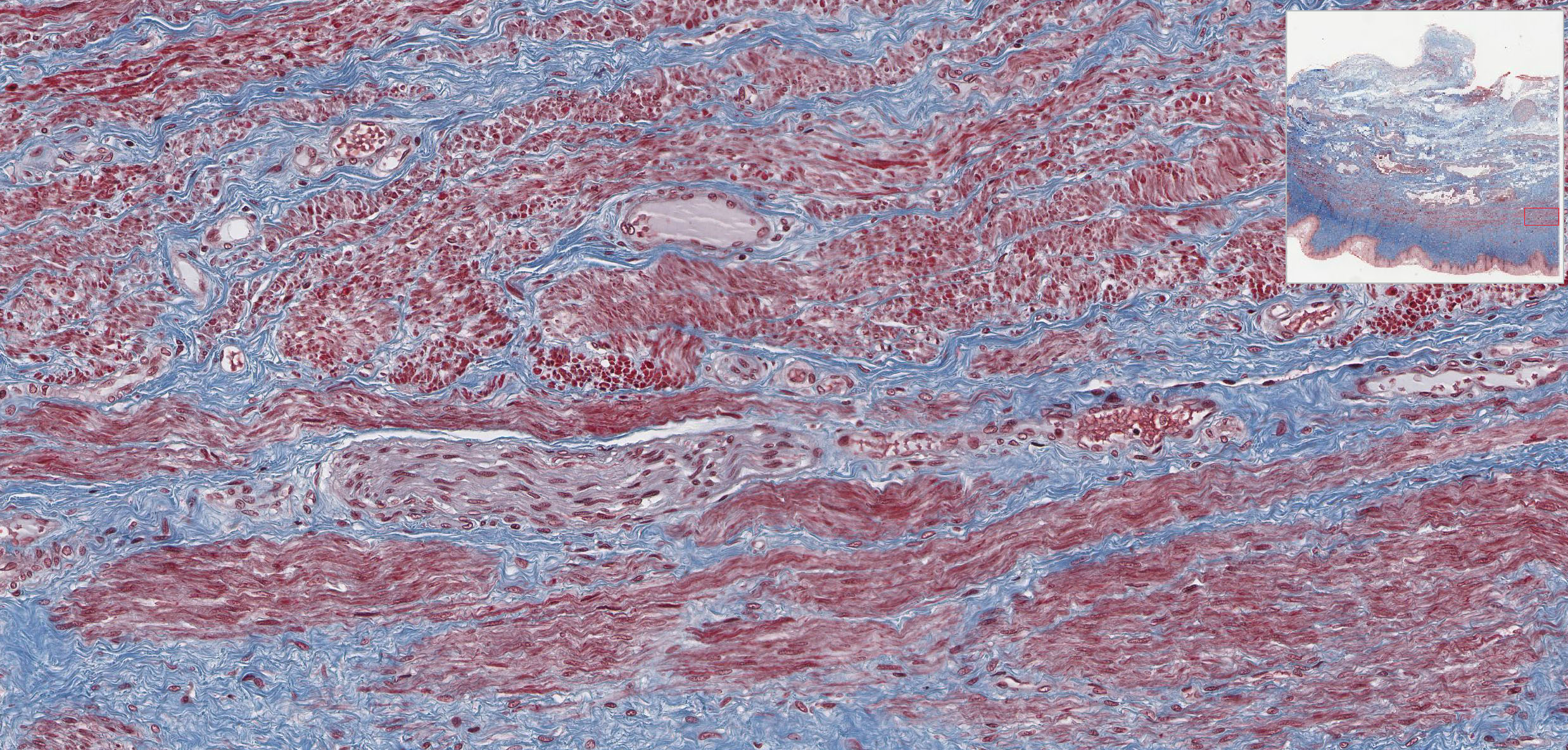
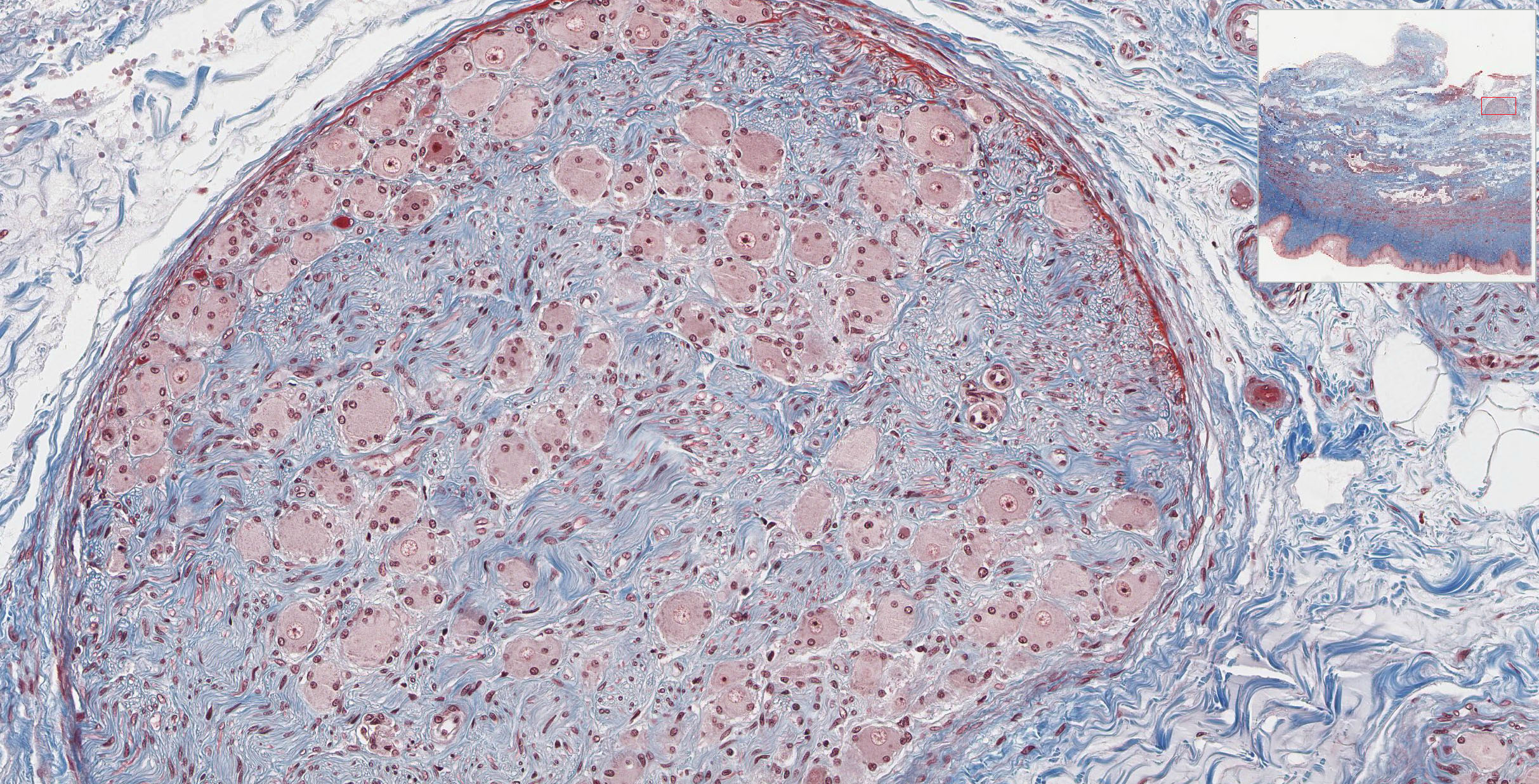
The connective tissue of the thick lamina propria View Image contains many elastic and collagen fibers throughout and many small veins in the deeper region. The subjacent smooth muscle layers View Image are arranged in poorly defined inner circular and predominant outer longitudinal layers; it may be easier to discern the smooth muscle from the surrounding connective tissue in the trichrome-stained section View Image. The connective tissue (seen particularly well in sections stained with trichrome) of the outer adventitial layer also contains many elastic fibers, thus contributing to the overall distensibility of this region. Also evident in the adventitia in both H&E View Image and trichrome-stained sections View Image are parasympathetic ganglia that innervate the erectile tissue (i.e. the numerous small veins) of the lamina propria.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Click on a question to reveal the answer.
In what way do the secretory cells of the uterus change at ovulation?
At ovulation, glycogen in the secretory cells of the uterus begins to move toward the basal end of the cell. Histologically, this is seen as cells with empty space at the base. This is called “basal vacuolization” and is an indication that ovulation has just taken place.
- Understand the structure of the chorionic villi and how it is changing with placental age.
- Be able to describe the delivery of nutrients and oxygen to the embryo based on the structure of the placenta.
Slide 253 Placenta, early, H&E View Virtual Slide
Slide 255 Placenta, late, 20x H&E View Virtual Slide
Slide 255 Placenta, late, 40x H&E View Virtual Slide
With the implantation of a zygote into the uterus, a number of changes take place in the endometrium. Cells from the embryonic trophoblast invade the uterine mucosa, secretions from these cells coalesce to form lacunae in the endometrium, which is now termed the decidua. These spaces also contain maternal blood. The trophoblast rapidly invades the decidua forming primary villi, that contain only trophoblast (outer syncytiotrophoblast and inner cytotrophoblast) cells. Mesenchyme and blood vessels form the core of the secondary villi. During the second half of pregnancy the cytotrophoblast cells disappear and the capillary basal lamina fuses with that of the syncytiotrophoblast to improve exchange.
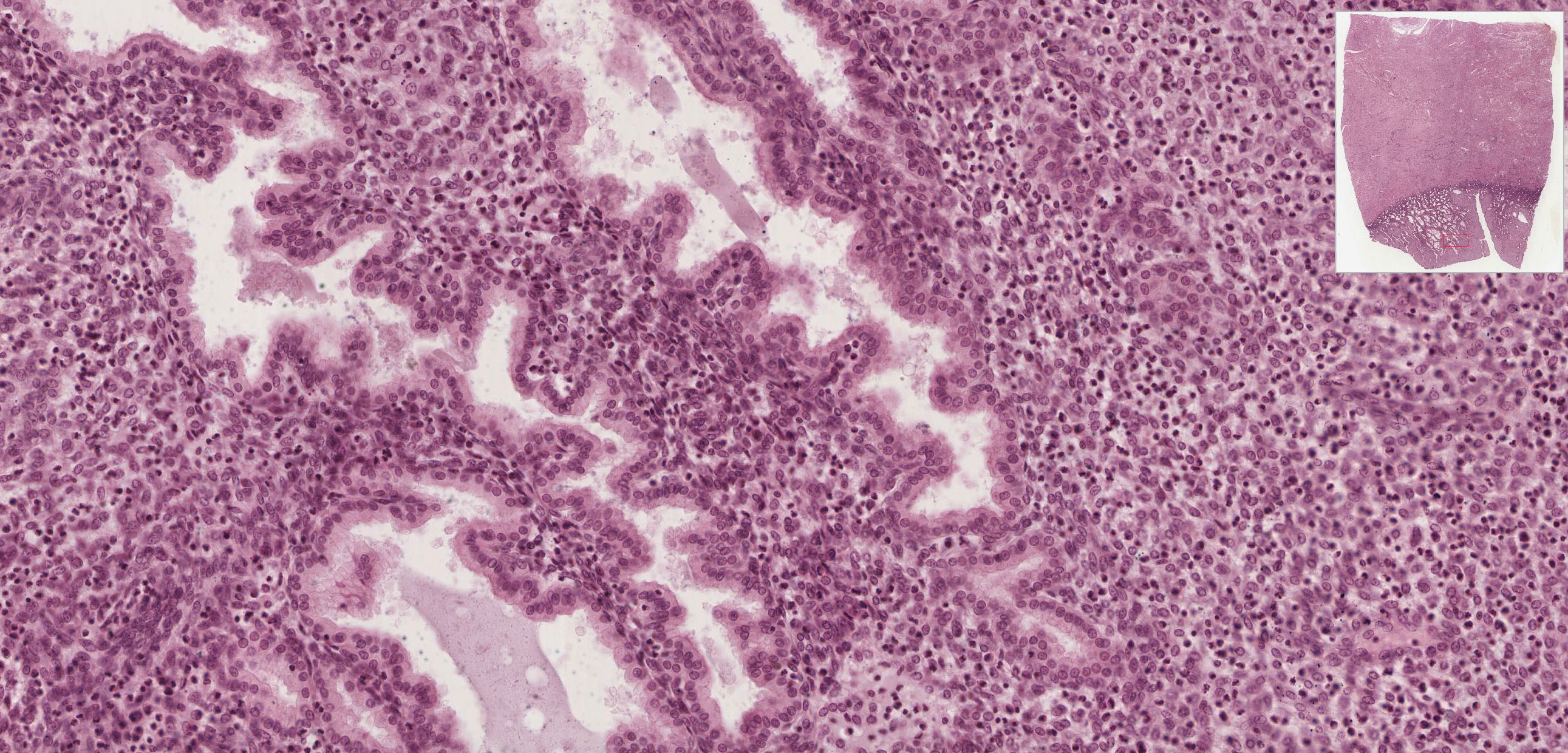
Begin with slide 253 View Virtual Slide, which is early placenta. The entire field is filled with villi cut predominantly in cross section. Mesenchymal tissue and fetal blood vessels (filled with fetal RBCs, which, in the early stages of fetal hematopoiesis, still have nuclei) make up the core of each villus which is covered by a trophectoderm shell View Image of inner cytotrophoblast and outer syncytiotrophoblast cells. The syncytiotrophoblast cells are more eosinophilic and have smaller nuclei. As the name implies, these cells result from the fusion of many cells and thus have many nuclei per cell and no discernable lateral boundaries. In contrast, cytotrophoblast cells are clearly demarcated, have a single, large nucleus and basophilic cytoplasm. Notice that there is almost a complete ring of cytotrophoblast cells covering each villus in early placenta --these cells are thought to function as stem cells for the formation of the syncytiotrophoblast, which is why they may disappear later in pregnancy.
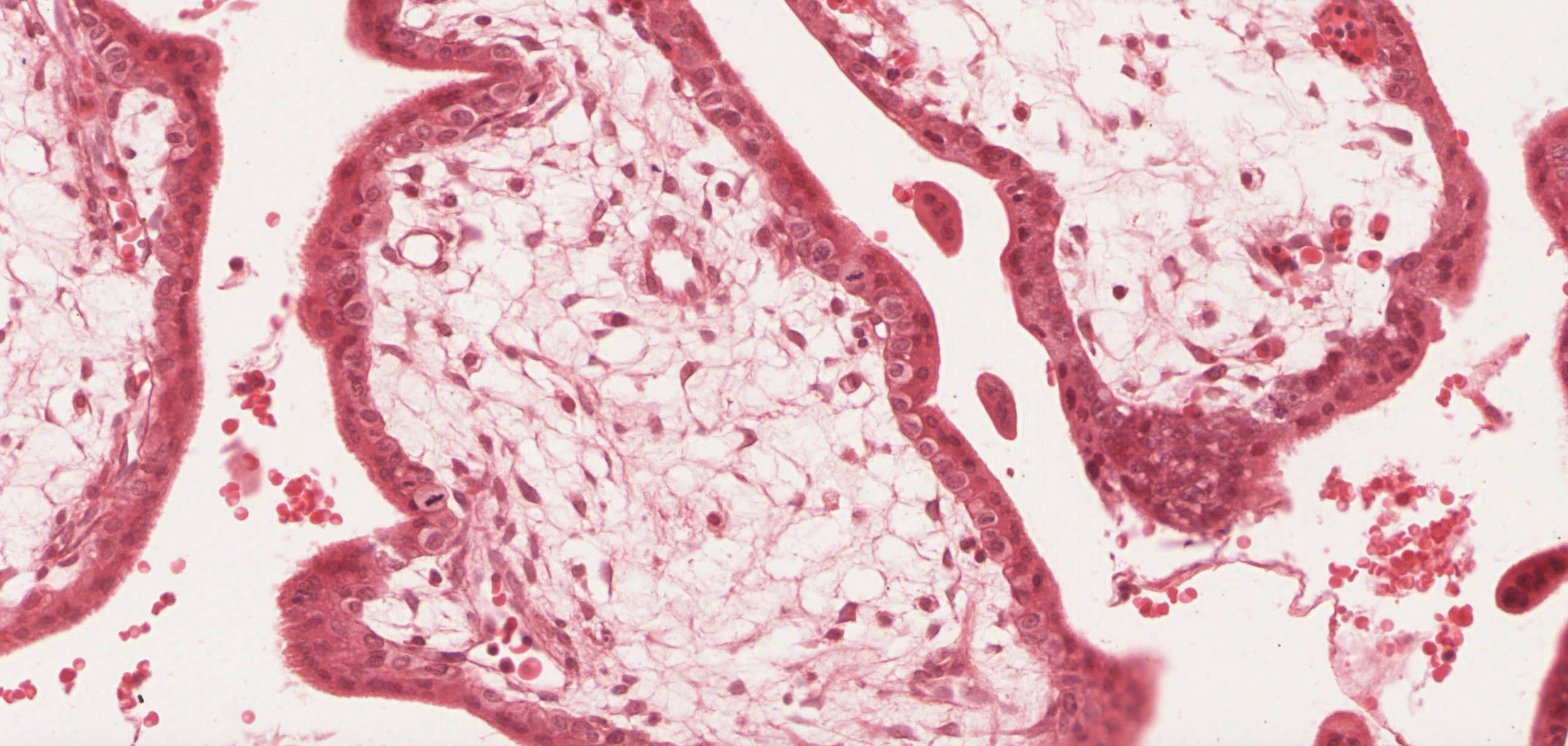{kind=link}
Study slide 255 View Virtual Slide, which is a late placenta. Although you will be able to see an occasional cytotrophoblast cell, you will not be able to see forming villi in this late placenta. Examine the trophectoderm shell View Image, which now consists of multinucleate syncytiotrophoblast cells located on the outer surface of the villi as well as an occasional residual underlying cytotrophoblast cell. The nuclei of the syncytiotrophoblast cells often form aggregates of so-called "syncytial knots" View Image that may look like specks of dirt when viewed at lower magnification. Syncytiotrophoblast cells that are bathed in maternal blood have apical surfaces specialized for absorption (microvilli), pinocytosis, and exocytosis and an appearance typical of secretory cells. Also examine the numerous macrophage-like (Hofbauer) cells View Image that can be found within the connective tissue of the placental villi. These cells are characteristically large, with elliptical, eccentric nuclei and extensively vacuolated cytoplasm. (FE5)
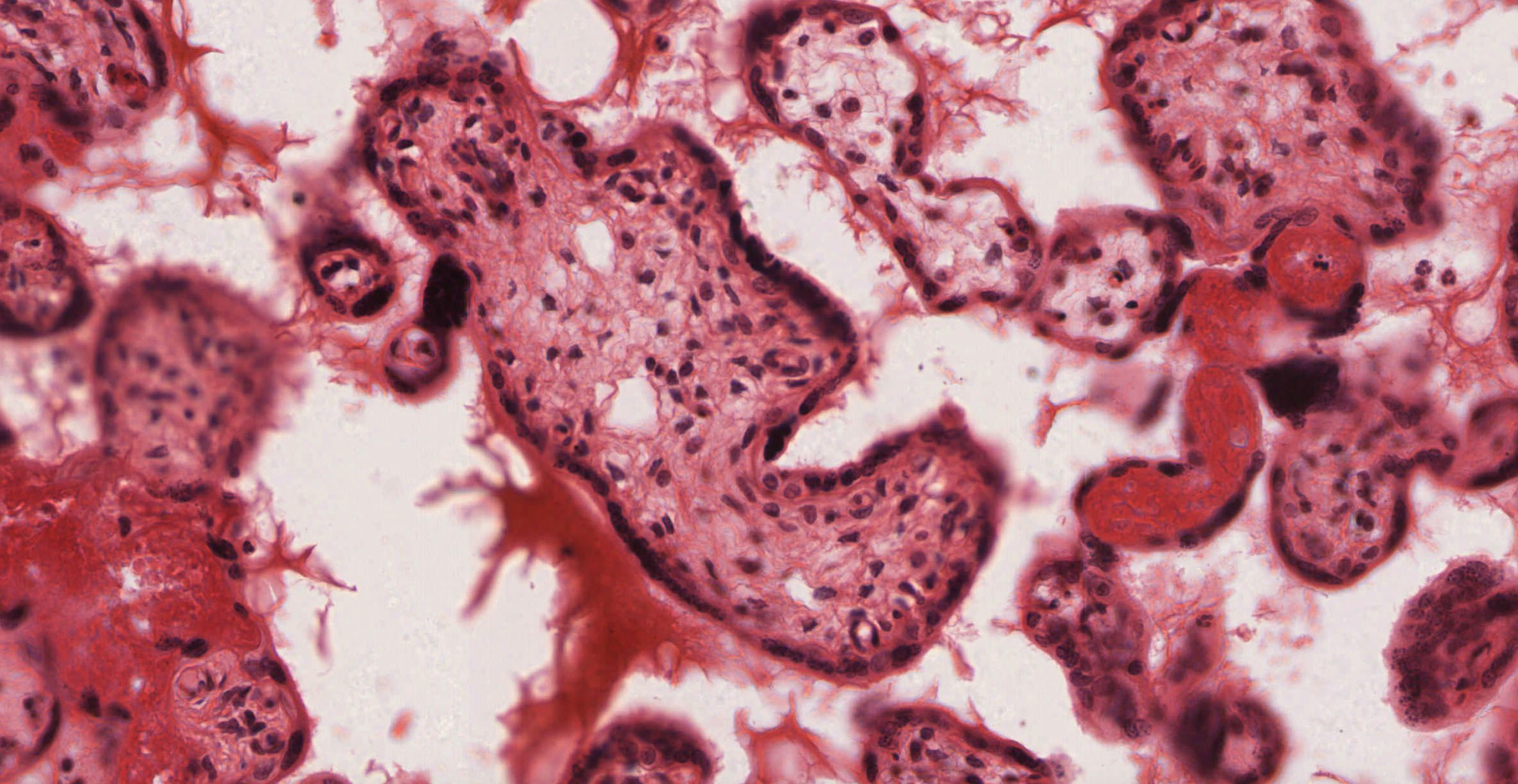{kind=link}
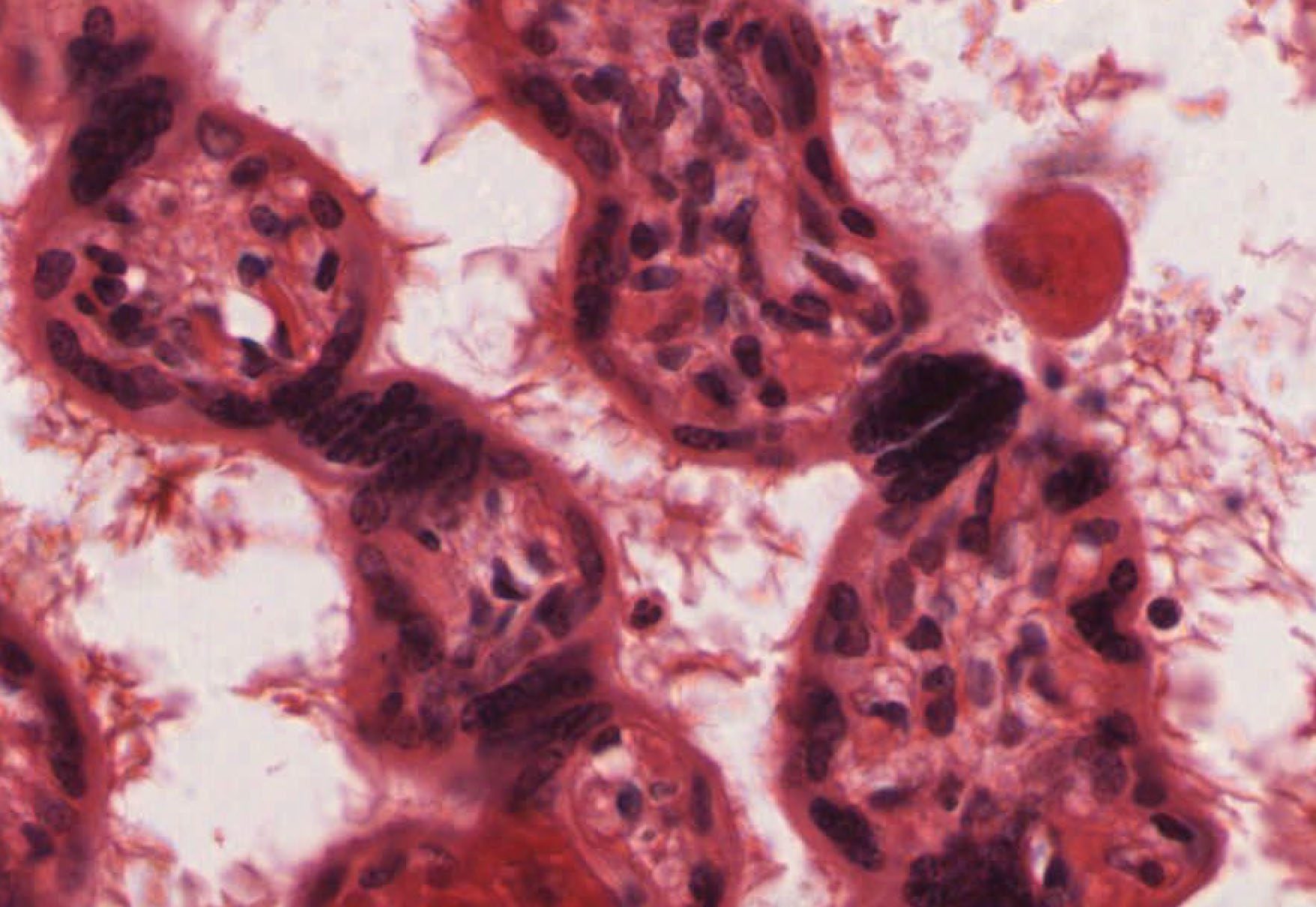{kind=link}
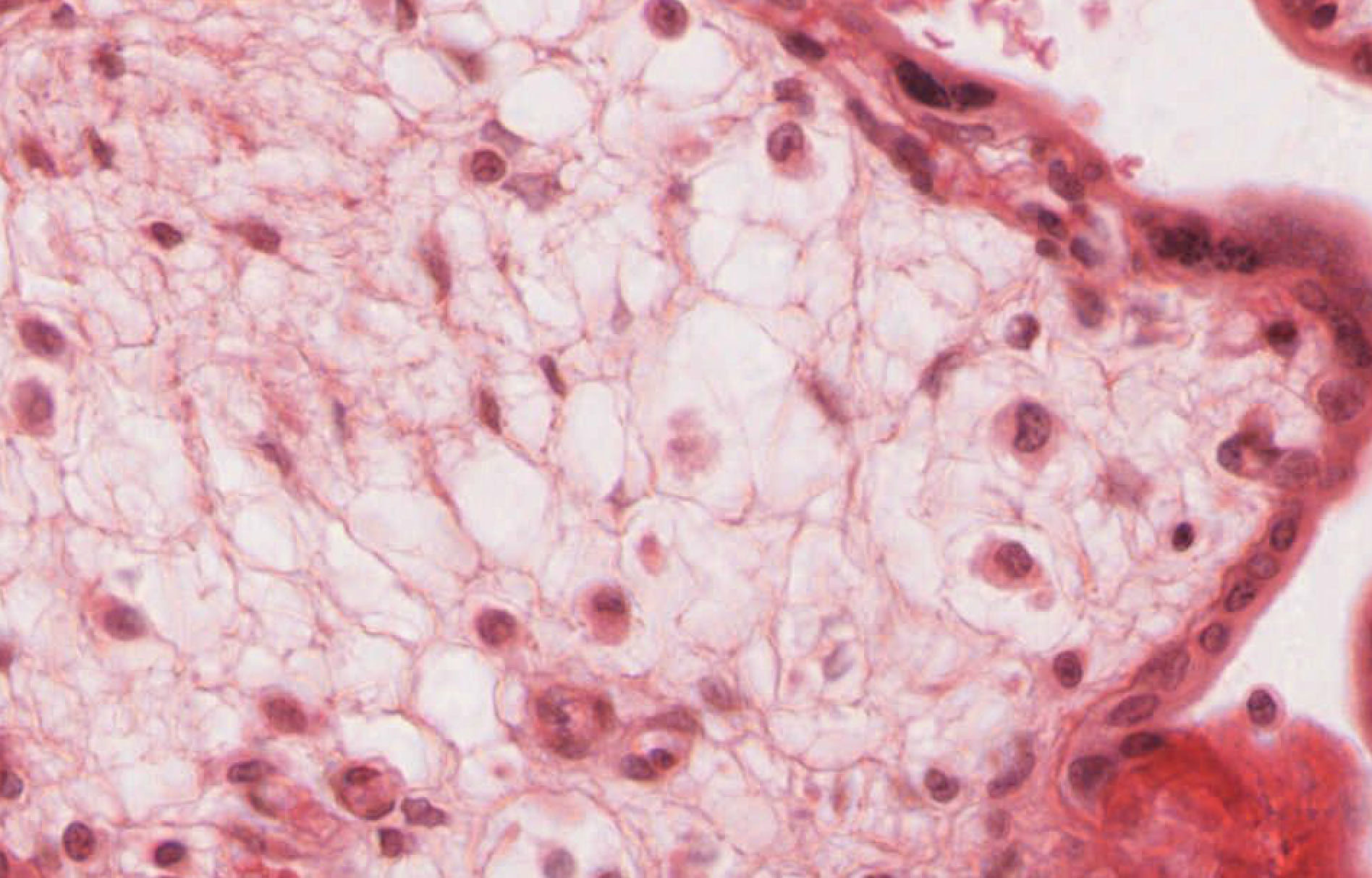{kind=link}
Large quantities of an amorphous eosinophilic substance, fibrinoid View Image, may also be deposited in the intervillus space in this older placenta. This material clings to the villus surface and may bind several villi together. The amount of fibrinoid in combination with the appearance of the connective tissue in the villi can be used to stage the placenta.
{kind=link}
289 Placenta - Placental Villus (mid pregnancy) View Virtual EM Slide
Examine the ultrastructure of the syncytiotrophoblast cells, particularly their external surface. Note the paucity of cytotrophoblast cells at this stage of development. Be certain that you know the components of the placental barrier, how they may change during pregnancy and why.
Click on question to reveal the answer.
What layers must nutrients cross to get from maternal to fetal blood in early and late pregnancy?
Early in pregnancy, nutrients from the maternal blood have to travel through the syncytiotrophoblast cells, a shared basement membrane, cytotrophoblast cells, the basement membrane of the capillary and finally the endothelial cell. As pregnancy progresses, the placenta changes. The cytotrophoblast is essentially non-existent and so nutrients cross the cells of the syncytiotrophoblast, its basement membrane, the basement membrane of the capillary and the endothelial cell.
1. Which of the following cell types commonly have prominent lipid droplets in their cytoplasm?
- Cells of the adrenal medulla
- Theca lutein cells
- Parathyroid chief cells
- Sertoli cells
- Thyroid parafollicular cells (C-cells)
- Acidophils of the anterior pituitary
Answer
Correct answer 2., theca lutein cells. These are steroid-secreting cells and they will have abundant smooth ER and lipid droplets in their cytoplasm.
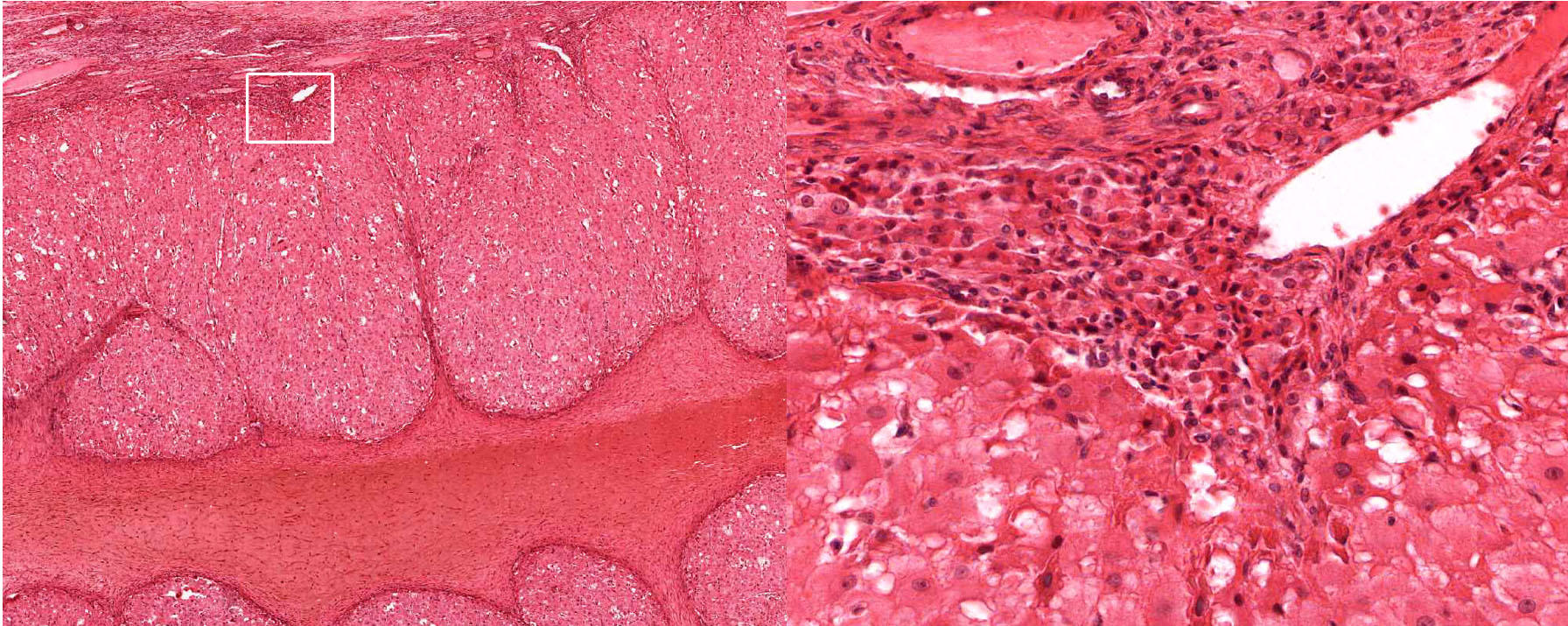
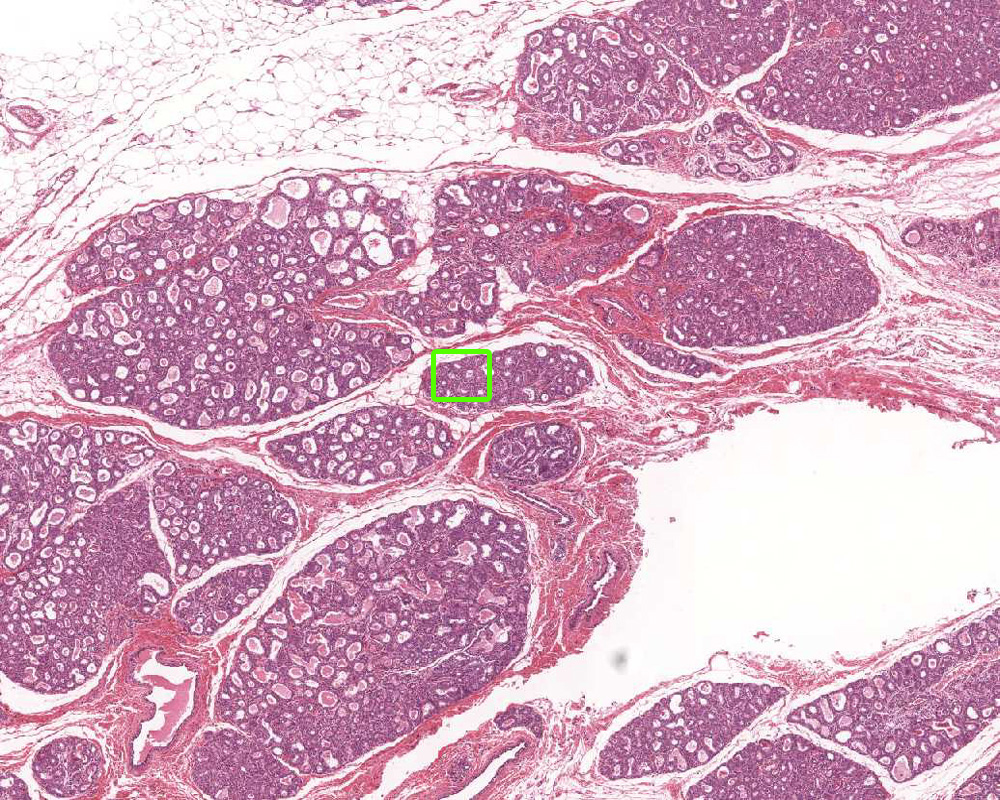
2. Identify the tissue/organ shown in this micrograph.
View Low Mag Image

View High Mag Image

- Normal thyroid gland
- Hypoactive thyroid gland
- Hyperactive thyroid gland
- Inactive mammary gland
- Active mammary gland
- Prostate gland
- Seminal vesicle
Answer
Correct answer 4., inactive mammary gland. Note the absence of secretory acini (ducts only).
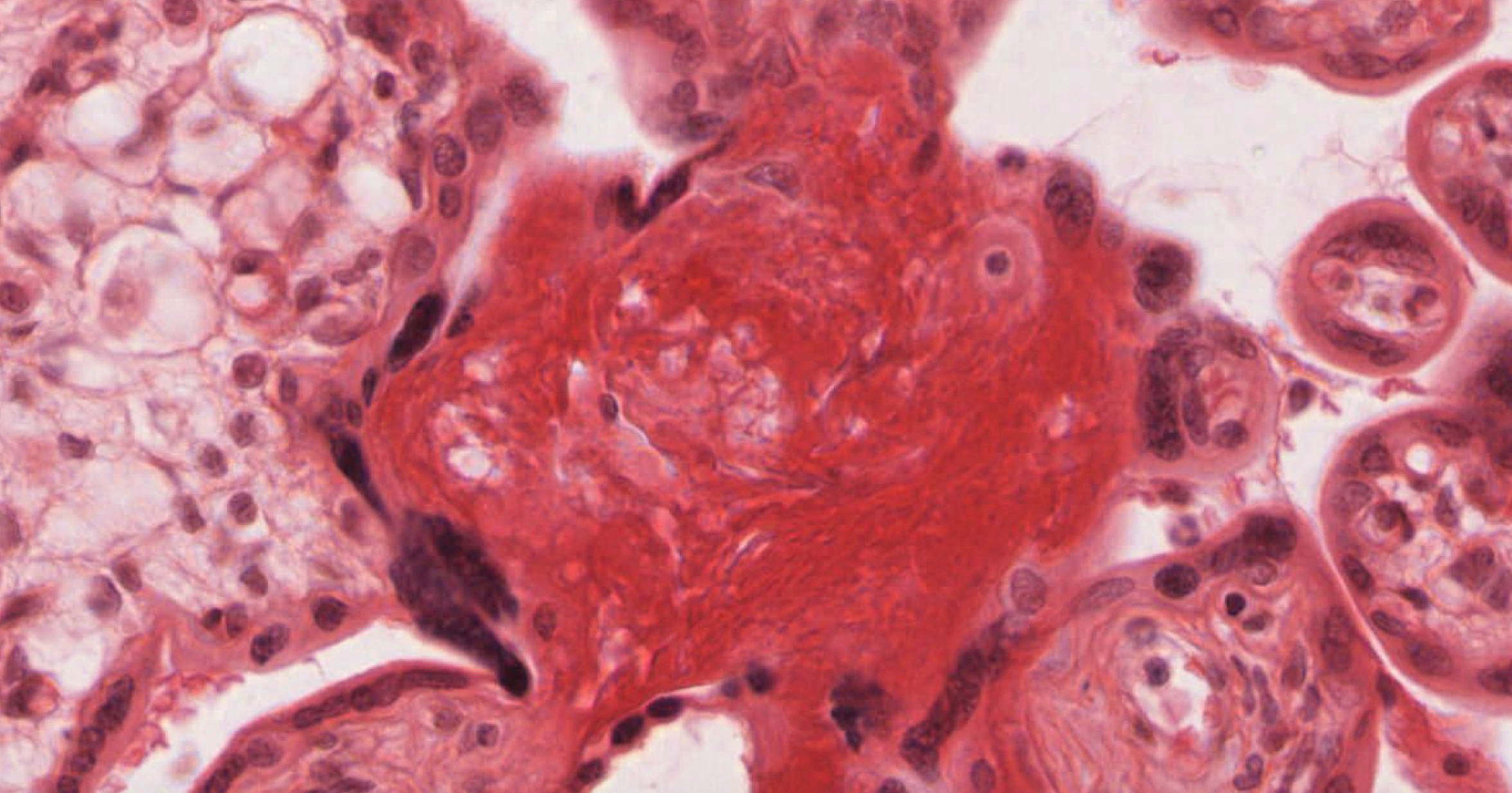
3. Which statement(s) is/are true about the cells marked in these micrographs?
View Image
- These cells produce LH.
- These cells produce FSH.
- These cells will develop into an atretic follicle.
- These cells will develop into a corpus albicans.
- ALL of the above statements are true.
Answer
Correct answer 4. The tissue shown is a CORPUS LUTEUM, which secretes (primarily) progesterone and will eventually develop into a corpus albicans.
4. Identify the product that is secreted by the cell indicated by the arrow in the high magnification image.
View Low Mag Image
View High Mag Image
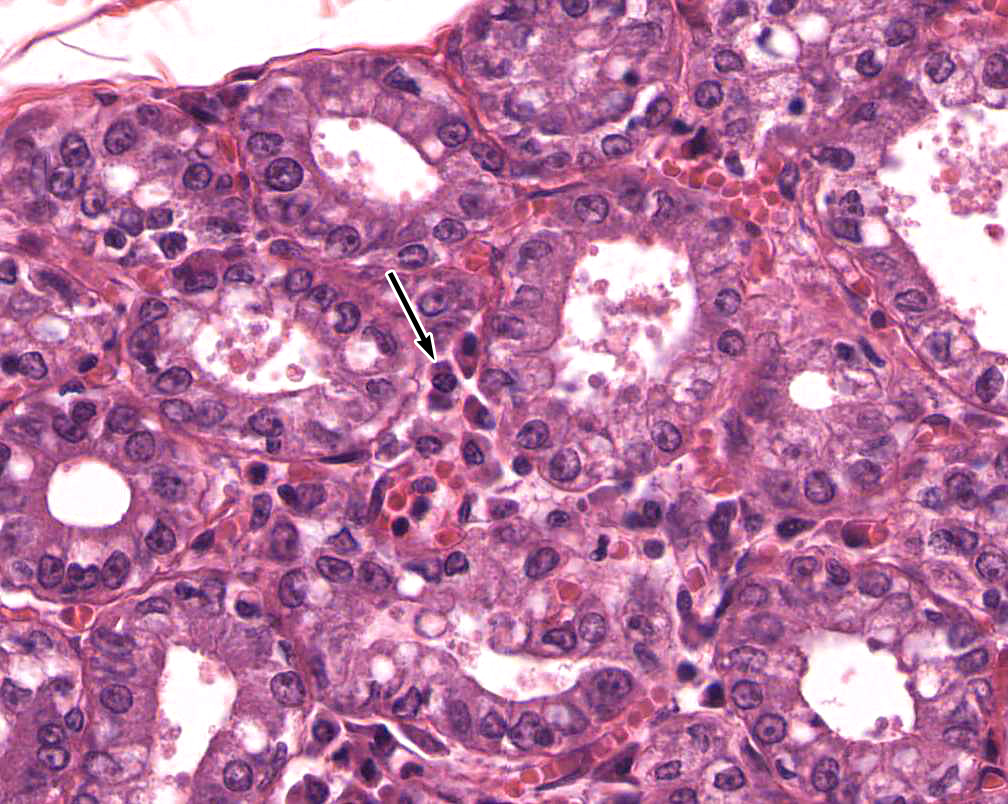
- Immunoglobulin A (IgA) dimers
- Calcitonin
- Thyroid hormone (T3 or T4)
- Testosterone
- Luteinizing hormone
- Prostate specific antigen
Answer
Correct answer 1., immunoglobulin A (IgA) dimers. The cell indicated is a plasma cell in the lamina propria of an active mammary gland. Plasma cells in glands produce stabliized dimers of IgA known as SECRETORY IgA that is then taken up and transported across the epithelium into the lumen of the gland.
5. Which statement(s) is/are true about the layer of cells indicated by the black arrow?
View Image
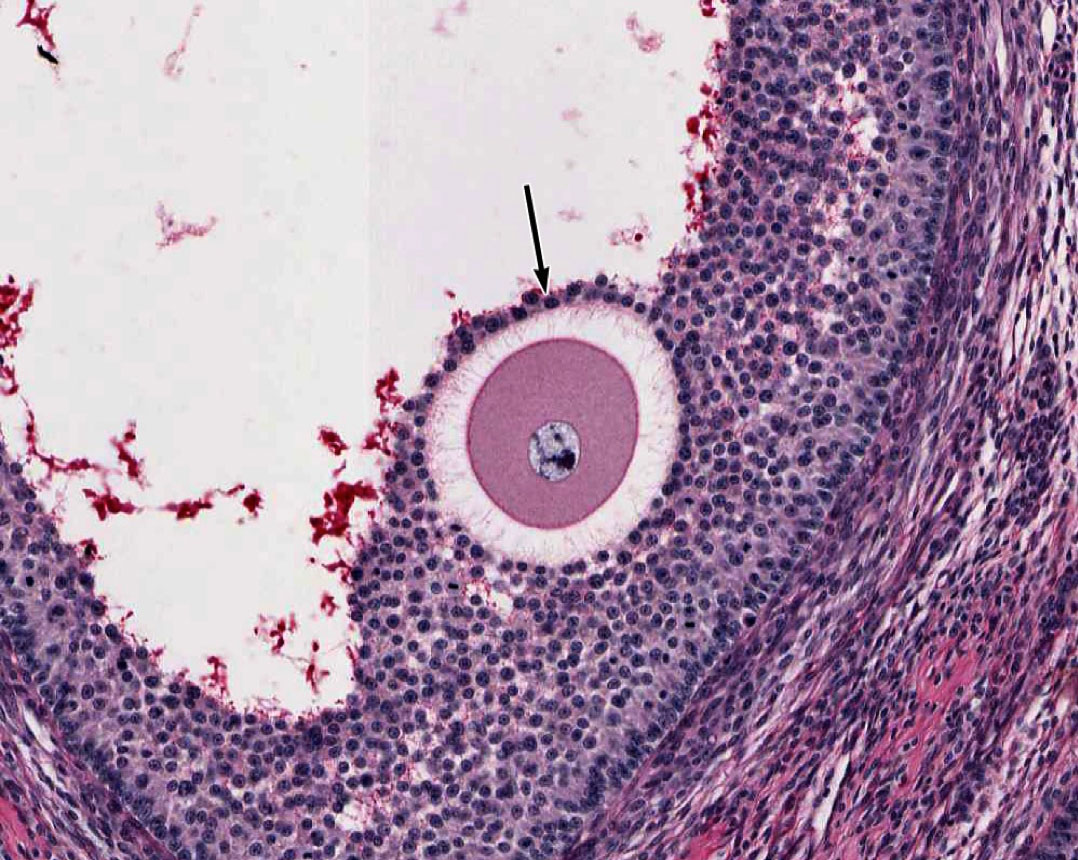
- These cells will convert into granulosa lutein cells after ovulation.
- These cells are derived from the theca interna.
- These cells are removed from the oocyte when ovulation occurs.
- These cells make contact with the oocyte via gap junctions
- These cells differentiate into trophoblast cells if fertilization and implantation occur.
Answer
Correct answer 4., they make contact with the oocyte via gap junctions. Cells of the corona radiata are ovulated together with the oocyte. They are derived from granulosa cells and make contact with the oocyte via gap junctions. The granulosa cells that are left in the follicle after ovulation convert into granulosa lutein cells.